
0 引言
联合国环境规划署2009年的报告中指出,红树林、滨海盐沼和海草床等滨海湿地生态系统能够捕获和长期储存大量有机碳,因而成为地球上最高效的碳汇之一,被称为滨海“蓝碳”[1]。这三类蓝碳生态系统的全球现有面积约为36~185万hm2,大约储存了全球90~327亿t的碳,相当于每年减少了1.4~4.6亿t的二氧化碳排放当量[2]。其中,红树林主要分布在热带和亚热带地区,滨海盐沼主要分布在温带地区,海草床则从极地到热带均有分布[2-3]。这三大生态系统除了固碳增汇的功能以外,还具有消波减浪、促淤护岸、生态服务等多种功能,可有效缓解全球气候变化和海平面上升带来的负面影响,对于增进人类福祉和维持全球生物多样性具有重大意义。
碳库(carbon pool)是沉积物、植被(指植物群落)、水体和大气等碳储存载体,可以吸收和释放碳。根据国际上已有的滨海蓝碳生态系统监测指南[4-5]中的定义,三大滨海蓝碳生态系统主要由植被碳库和沉积物碳库(也叫土壤碳库)两大部分组成。其中,植物通过光合作用固定二氧化碳形成生物量,并通过根系在厌氧沉积物环境中的分解进一步形成沉积物中的有机碳。沉积物碳库根据来源可分为内源碳和外源碳,内源碳是由植物本身形成并进入沉积物中的碳,而外源碳则是来自于周边邻近的生态系统(如流域和近海)的碳,通过水体中沉积物输运,并被捕获、埋藏后加入沉积物碳库[6⇓-8]。植被碳库是由植物生长的固碳作用形成的,是大气二氧化碳被捕获的主要媒介,这个碳库相对来说较为活跃。沉积物中的有机碳可长期封存,时间尺度达几百到上千年,因此沉积物碳库相对植被碳库来说容量更大,更加稳定,是有机碳的长期归宿。根据全球尺度的初步估算,滨海蓝碳生态系统的沉积物碳库大约每年固定4 500~21 600 万t有机碳,贡献了全球海洋沉积物碳库增量的30%~50%[9⇓-11]。由于滨海植被碳库和沉积物碳库储碳能力巨大,因此,当滨海湿地遭到破坏时,这两大碳库会产生显著的碳排放,成为重要的碳排放源。根据估算,全球每年由于滨海湿地的破坏或者退化,会造成1.5~10.2亿t二氧化碳从滨海湿地碳库中被释放出来[3]。因此,在滨海蓝碳生态系统的碳汇机理研究中,需要特别关注植被碳库和沉积物碳库这两大碳库的变化。
事实上,滨海蓝碳生态系统碳库内部不同组分之间和不同类型碳库之间存在动态的相互作用,共同造成了碳库整体的变化。虽然这种碳库的相互影响还未被系统性地梳理和讨论,但是对于理解滨海蓝碳系统的固碳过程和能力具有重要的意义。基于以往的研究,本文试图通过梳理文献来阐述滨海蓝碳生态系统碳库间的相互作用及其对系统碳汇能力的影响。综述将从以下三个角度展开:滨海湿地植被碳库内部的相互影响、滨海湿地沉积物碳库内部的相互影响以及植被碳库和沉积物碳库之间的相互影响。基于这三个角度的分析,总结蓝碳生态系统碳库研究的现状和问题,提出对未来研究的展望。
1 植被碳库之间的相互作用
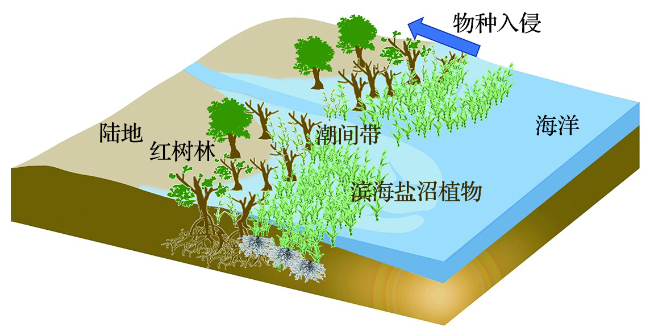
红树林是热带、亚热带海岸带的一种高生产力生态系统,也是热带地区碳储量最丰富的森林之一[13]。中国红树林具有“三高”特性,其中的“高生产力”特指红树林储碳和固碳的高效性[14]。从碳储量来看,在全球尺度上红树林地上生物量碳密度为6.6~250.8 Mg C·hm-2(生物量为16.6~627.0 Mg·hm-2),具有很高的生产力[15]。以草本植物为主的滨海盐沼和海草床生态系统,其单位面积植被碳库总量均小于以木本植物为主的红树林生态系统。在我国,由于滨海盐沼的分布面积最广,其整体的储碳和固碳能力也最强,特别是在高潮带滨海盐沼中,地上生物量占总碳库的比重较大,具有重要的固碳作用[16]。海草床是全球性的生态系统,在除南极洲以外各大洲均有分布,具有很高的初级生产力[10,17-18]。有研究通过数据整合计算出全球海草生物量碳密度为0.61~7.29 Mg C·hm-2,证实海草床是不可忽视的植被碳库[18]。
通常来说,红树林分布在热带到亚热带地区,而滨海盐沼则广泛分布在热带到寒带,这两种植被类型均可以在亚热带潮间带形成生态群落[19]。如在中国、澳大利亚和美国等地的海岸带形成了红树林和滨海盐沼沿纬度分布的生态位重叠[19-20]。在气候变化的背景下,在澳大利亚和美国的一些沿海地区,红树林的分布呈现出极向扩张并入侵到滨海盐沼中,尤其在美国,这种红树林入侵盐沼的群落演替增加了滨海盐沼的生物量碳储存[21-22]。在我国的滨海湿地,互花米草快速入侵红树林和乡土盐沼,这种植被碳库之间的相互影响与发生在美国和澳大利亚等地的情景相比存在一定的差异性 [12]。互花米草兼具有性繁殖和无性繁殖,其植株个体定植在潮滩后,通过无性繁殖迅速形成圆形的克隆斑块“精灵圈”[23],当相邻斑块扩张融合后,形成均匀连片的植被。互花米草在潮间带的分布呈现“单峰型”曲线,中潮带是最佳生长区,也是红树林-互花米草种间竞争最激烈的潮位[24-25]。在福建漳江口中潮位区域,互花米草侵占红树林前缘光滩,形成典型的红树林-互花米草 (“树-草”) 交错格局,原先提供红树植物幼苗扩散定居的前缘光滩也逐渐转化为典型的“树-草”竞争区域[20,25]。
基于遥感图像分析,我国互花米草盐沼的总面积从2007年的34 451 hm2[26]增加到2015年的54 580 hm2[27]。互花米草通过在我国沿海潮滩上的迅速扩张,贡献了额外的蓝碳。根据LIU et al[28]的盐沼地下碳密度和刘钰 等[29]的地上生物量碳密度计算,由互花米草生物量增加而增加的碳密度为80.41 Mg C·hm-2。据此,CHEN et al[12]估计,在2007年至2015年期间,互花米草在全国贡献了1.62 Tg C的蓝碳储量。因为各生态系统不同的生物量和结构,以全球平均水平而言,红树林植被碳库比滨海盐沼植被碳库储存了更多的碳[30]。然而,在互花米草入侵红树林的过程中,尽管互花米草的地上生物量比红树植物要低,但是由于互花米草的地下生物量占其自身碳库的比重大,根系生物量增长迅速,植被碳库的总量有可能增加。根据对漳江口“树-草”交错区的研究,当红树林被互花米草占据之后,植被碳库的碳储量密度由原先的168 Mg C·hm-2上升为1 948 Mg C·hm-2[12]。
2 沉积物碳库之间的相互作用
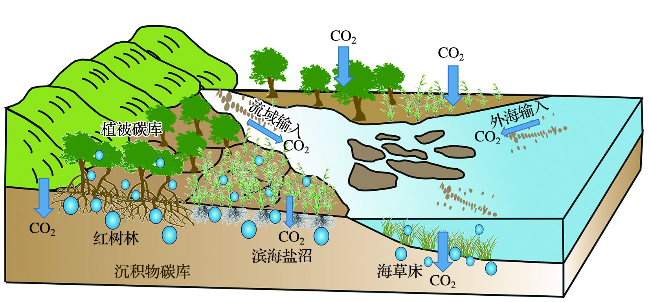
沉积物碳库是蓝碳生态系统中最主要的碳库,碳储量通常占到红树林生态系统总碳储量的50%~90%,在滨海盐沼和海草床生态系统中占比甚至超过98%[13,18,33-34]。沉积物碳库的有机碳埋藏过程是将地表活跃的碳库最终转化为长期稳定的地质碳库的关键步骤。近期已有大量研究关注全球尺度下的蓝碳生态系统沉积物碳库碳储量和年际增量的问题[9,11,35],并指出了沉积物碳库在响应气候变化方面的重要贡献。事实上,虽然地表生态系统光合作用十分活跃,但是由光合作用固定的有机碳最终只有大约0.5%进入到海洋沉积物之中被长期保存,因此,从长时间尺度看,沉积物碳库整体保存率较低[36]。相对于大洋生态系统来说,由于滨海湿地蓝碳生态系统发育在潮滩系统之上,会受到径流输入、外海输入、潮汐波浪作用、沉积物输运、地下水循环等复杂过程的影响[37],其有机碳的保存过程会更加复杂。
为了进一步提高对沉积物碳库碳汇的认识,除了观测沉积物碳库的储量和增量以外,还需要进一步理解沉积物碳库的成因和控制因素,其中有机质来源和碳埋藏过程是理解沉积物碳库变化的关键科学问题。对于蓝碳生态系统的沉积物碳库来说,内源碳和外源碳共同控制了沉积物碳库的碳汇能力,但两者的比例在不同蓝碳系统中会出现相当大的差异。海草床的沉积物中大约有50%的碳是外源碳[8],相比之下,红树林和滨海盐沼系统中的内源碳占比较高[7],但是研究发现外源碳有时也可以在红树林、滨海盐沼的沉积物碳库中占据主导地位[38]。与此同时,蓝碳生态系统中的沉积物碳库也可以作为输出源,通过水体的动力学交换输出到周边其他生态系统之中,即沉积物碳库的“输入(inwelling)”和“输出(outwelling)”问题[38]。因此,从更大尺度来看,蓝碳生态系统的沉积物碳库是本地沉积物碳库和周边生态系统沉积物碳库之间相互作用的产物(图2)。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
为了厘清沉积物碳库之间的相互作用,需要开展沉积物有机碳溯源研究和水-沉积物界面通量变化研究。在这些研究中,“端元分析”和“箱式模型”发挥了非常重要的作用。“端元分析”是一种建立在物质守恒原理之上的物源追踪方法,包括示踪标记物的确定和物质混合模型的建立[39-40]。示踪标记物目前主要采用碳氮比(C/N)、稳定同位素(如δ13C、15N)以及生物标志物,并且更多的研究是将多种示踪标记物进行联合应用[41-42]。蓝碳生态系统的沉积物有机碳来源“端元分析”可以采用“二元混合模型”,即只区分内源碳和外源碳[38,43⇓-45];也可以采用“三元混合模型”,即本地植物产生的有机碳、流域输入的有机碳和外海输入的有机碳这三个端元[41]。
“箱式模型”是环境领域进行大气环境容量估算的常用工具。这种模型将研究的空间范围看成是一个尺寸固定、状态稳定的“箱子”,通过输入与输出来计算各个箱子之间的物质交换[46]。在解决海洋学的复杂界面问题时,“箱式模型”能化繁为简,在碳循环领域中发挥着重要作用[47]。近期,基于“箱式模型”,已经开发了用于沉积物碳平衡评估的ICBM模型(introductory carbon balance model)及其衍生模型,可覆盖不同类型滨海蓝碳生态系统的温室气体排放模式[48-49]。在滨海蓝碳生态系统中,“箱式模型”可以帮我们更好地理解沉积物碳库之间的交互作用:基于端元分析,沉积物碳库的有机碳输入主要由上游流域沉积物碳库输入、外海沉积物碳库输入和本地生物输入组成,输出主要来自沉积物碳库向外海输出和沉积物内的生物呼吸作用;若只考虑净效应,则只需要考虑流域输入、外海净输入(或输出)以及本地生物的净输入,最终以碳埋藏速率作量化表征。因此,沉积物碳库之间的交互作用是一种系统性、可远距离发生的作用,取决于流域输入和来自外海的物质交换。
全世界每年有大约100~200亿m3的泥沙从流域输送到海洋[50],其中亚洲的贡献最大[51]。在我国,根据大多数文献的沿用数据,历史上黄河的输沙量最高,达到每年1.2亿t,而长江的年输沙量约为0.5亿t[40]。这些流域输入物质大部分都会堆积在河口三角洲区域,使得发育在这些区域的滨海湿地具有较高的沉积速率,并随沉积发生较显著的碳埋藏过程。陆地沉积物碳库在这些河口地区对滨海湿地沉积物碳库产生重要的影响。黄河口的滨海盐沼沉积物碳库以陆源有机碳和植物有机碳为主,海源有机碳含量较低[52]。黄河口滨海湿地的放射性元素测年结果显示其长时间尺度的沉积速率达到1.0~2.8 cm·a-1,沉积物有机碳埋藏速率为65~193 g C·m-2·a-1[16,53]。长江口及其毗邻的杭州湾潮间带沉积物的有机碳主要来源于长江,陆源有机碳贡献超过55%[44,54-55]。长江口滨海盐沼的沉积速率为1.1~4.9 cm·a-1,其碳埋藏速率为73~485 g C·m-2·a-1[16],其毗邻的杭州湾南岸滨海盐沼的长期沉积速率达到了2.1~2.8 cm·a-1,沉积物碳埋藏速率为119~159 g C·m-2·a-1[44]。值得注意的是,由于上游流域下垫面的变化和水坝的建设,自1950年以来,黄河的输沙量以每年1.7%的速度递减,长江的输沙量也在以每年0.4%的速度下降[56],这就意味着将来流域沉积物碳库和滨海湿地沉积物碳库之间的作用会减弱。
与此同时,通过水动力的混合过程,海源颗粒物也能被潮汐和波浪带入潮间带。当流域输入的沉积物减少时,外海的输入贡献有可能增强。通过对我国南方河口地区的现场观测和“箱式模型”分析,CHEN et al[57]发现由于水电站建设造成流域泥沙减少后,来自外海的沉积物输运会被加强,而且在这个过程中,河口内发育的滨海湿地会起到非常重要的调节作用。因此,考虑到未来流域减沙和海平面上升的背景,外海沉积物碳库和滨海湿地蓝碳生态系统沉积物碳库之间的交互作用有可能会进一步加强。
3 植被碳库与沉积物碳库之间的相互影响
在植物冠层捕获颗粒物的同时,植物的地下部分也对沉积物碳库产生重要的影响。滨海湿地植被具有复杂的三维根系结构,根系特征及其分布深度取决于植物的类型。红树林的根系较为复杂,既有地上部分的气生根,又有地下部分的细根。其中,地上部分的气生根和植物冠层共同引起对沉积动力学过程的扰动;而地下根系则可以通过根区膨胀作用,进一步抬升地表高程[80-81]。从全球多个地点的观测结果来看,不同类型的红树林根系平均额外贡献了75%的地表高程抬升,滨海盐沼根系则贡献了67%[53]。这种根区膨胀作用使得沉积物碳库得到了扩容,促进了此类碳库的发育。相比之下,蓝碳生态系统的草本植物产生根区膨胀主要以地下根系为主,除了主根之外,还包括大量直径小于100 μm的细根和须状根[32]。以滨海盐沼为例,大部分活根集中分布在大约30 cm的表层沉积区,但部分根系可以向下延伸超过1 m[32,44]。根系通过绑定、黏附沉积物颗粒,显著提高了表层沉积物的抗侵蚀能力,使得沉积物碳库更加稳定,促进了有机碳的埋藏[82-83]。
在全球海平面上升的背景下,淹水时间延长和淹水频率增加,是蓝碳生态系统面临的最重要胁迫之一[81]。由于淹水而造成的缺氧是影响植物生长的主要原因[84]。已有研究发现,潮汐可以通过周期性淹水直接作用于植被,持续淹水对植物的生长具有抑制作用,而短时间淹水则具有一定的促进作用[85⇓-87]。淹水条件通常由滩面高程和水位条件共同决定,这就形成了沉积物碳库影响植被碳库的主要机制:沉积物碳库通过滩面高程的变化来调节淹水过程,最终影响植物的生长。控制实验发现,在沉积作用为主的地区,适当的沉积速率可以促进互花米草、碱蓬等潮滩植物的生长,但是过高的沉积速率则会引起根系缺氧和硫化物累积,导致植株死亡[88-89]。在以侵蚀作用为主的地区,由于底部沉积物被侵蚀,植物会发生死亡,但是不同植物种类之间具有一定的差异[90]。因此,在相对海平面上升的背景下,一定程度上的滩面淤高能减少淹没时间、淹没频率和淹没深度,使得物理环境有利于蓝碳生态系统内植物的生长,维持整个生态系统的稳定性,形成沉积物碳库对植被碳库的正向反馈[91-92]。
通过植物生长和沉积动力地貌过程之间的交互作用,滨海蓝碳生态系统的植被碳库和沉积物碳库形成了有效的相互作用,即植被碳库的增长促进沉积物碳库的稳定性,而沉积物碳库的增长使得植被碳库能更好地应对海平面上升等胁迫,形成正向的反馈,使得系统保持稳定,具有应对气候变化的能力。
4 总结与展望
滨海蓝碳生态系统植物的竞争和演替会带来植被碳库之间的交互作用。外来物种互花米草的扩散已经被证明会显著影响我国蓝碳生态系统的碳汇能力。在我国南方的潮滩上,除了互花米草以外,外来红树植物(如无瓣海桑)也呈现扩散趋势,与本地种形成竞争,对蓝碳生态系统的碳汇功能造成了影响。20世纪80年代以来,我国部分红树林保护区开展了外来种的引种工作。自1985年以来,大约有6种外来红树植物被引种到我国南方的潮间带,包括来自孟加拉国的无瓣海桑、墨西哥的阿吉木、美洲的大红树和拉关木等[81]。由于对环境的高适应性和速生的特点,无瓣海桑和拉关木被广泛应用于南方一些困难立地条件下的红树林修复工作中,其中无瓣海桑已经成为我国红树林造林的主要物种,占据我国人工红树林面积的50%以上[93]。尽管无瓣海桑是否属于入侵植物尚存在较大的争议[94],但是外来红树植物对于本地红树林植物碳库的演化必然具有重要的影响,亟需开展进一步研究来厘清和预测外来植物带来的蓝碳生态系统碳汇功能的变化,为后续的生态修复工作提供指导。
沉积物碳库之间的相互作用主要体现在外源碳的输入和沉积过程中。在蓝碳研究领域,相比植被碳库,关于沉积物碳库的研究相对较少,其中较早的研究开展于上世纪80年代左右[4,95]。事实上,沉积物有机碳研究在海洋地质学和地球化学领域具有悠久的历史,最早可以追溯到1915年发表在Report of Danish Biological Station上关于海洋泥质沉积物中腐殖质有机碳含量的论文[96]。沉积物中有机质的来源和埋藏过程一直是海洋地质学和地球化学重要的研究内容,很多知识和方法可以进一步应用到蓝碳研究之中。例如,有机碳在潮间带的埋藏过程和物质搬运过程有非常密切的联系,是沉积物某一特定组分的搬运和沉积过程。因此,可以结合沉积学的传统物源分析方法,如沉积物组分、粒径、黏土矿物分析[97-98]以及地球化学的源汇示踪手段[99],来共同揭示有机碳的来源和去向问题。目前,在蓝碳沉积物碳库的碳储量和增量的观测上,主要观测参数集中在沉积物容重和有机碳含量,对于其沉积历史尚未开展系统性的研究。在这方面,可以充分利用碳库调查中采集的柱状样,同步开展沉积学的分析,对历史沉积环境和有机碳埋藏事件进行重建,以便更好地理解地质时期碳埋藏的过程和长时间尺度的沉积物碳埋藏潜力。
植被碳库和沉积物碳库之间的交互作用,是滨海湿地生物地貌学的重要研究内容[63]。生物地貌学主要研究生物对沉积动力条件和地貌过程的影响机制,以及沉积动力条件和地貌过程对生物发育的反馈机制[100]。对于以大型潮滩植物群落为特征的蓝碳生态系统,非常适合用生物地貌学手段来开展植被碳库和沉积物碳库之间交互作用的研究[101]。在这方面,目前至少有两个重要的问题有待进一步研究。首先是碳汇的时空尺度问题。植被的生长周期、沉积物的输运周期和碳埋藏的时间尺度并不相等。植被的空间分布覆盖了斑块尺度到景观尺度,沉积物的输运则涉及“流域-河口-近海”这样从源到汇的空间尺度,这种跨尺度的碳汇效应如何进行协同评估,是一个重要的挑战。其次是碳库潜力的阈值问题。从植被碳库来看,植物群落的发育和固碳能力具有一定的上限,例如红树林生态系统的固碳能力在其生长20~80年达到峰值[82],其后植被碳库难以继续提升。类似的,对于沉积物碳库来说,随着潮滩地形达到均衡态[102],沉积速率将逐渐下降。在两者交互作用中,沉积速率对于植被生长的促进作用也只发生在一定的阈值之内。因此,需要通过发展生物地貌学的观测技术和模拟能力来进一步探索和预测滨海湿地碳库的潜力阈值问题,为后续的滨海湿地生态修复和蓝碳管理提供支撑。
致谢
厦门大学顾肖璇、河海大学刘兵和浙江大学张子严协助完成本文的图件绘制和编辑工作,在此一并致谢!