
0 引言
作为蓝碳生态系统的重要组成部分,海草床已被大量研究证明具有很强的储碳能力,因此它是一种重要的、基于自然的缓解气候变化途径[1]。海草床沉积物有机碳库主要由自身光合作用形成的内源碳和通过捕获并截留水体悬浮物所构成的外源碳两部分构成[2]。相比于有机碳库,目前关于海草床沉积物中无机碳库的形成过程与埋藏通量的研究相对缺乏。在具有高钙化生物丰度的海草床中,沉积物无机碳储量可能超过有机碳储量,而沉积无机碳形成过程中伴随的CO2释放,可显著抵消有机碳埋藏所产生的碳汇效应[3-4]。具体而言,每生成1 mol的CaCO3,将向大气释放约0.6 mol的CO2[5]。因此,在评估海草床的“蓝碳”功能时,有必要同时考虑沉积物有机碳库与无机碳库的相对规模。开展海草床沉积物无机碳库研究,对于全面理解海草床在区域乃至全球碳循环中的作用具有重要意义。
海草床被视为全球无机碳埋藏的热点区域之一[6]。其生态系统中无机碳(可视为全部为CaCO3)的主要来源可分为外源输入与原位生产两类。外源输入的无机碳主要指来自于邻近生态系统的无机碳沉降,例如珊瑚礁生态系统及陆源生态系统中的碳酸钙颗粒随水体流动进入海草床生态系统,被海草冠层捕获并埋藏于沉积物中[1]。原位生产的无机碳则主要源于生活在海草床中的钙化附生藻类和钙化底栖生物。此外,有研究表明部分海草物种(如泰来草)自身具备生物钙化能力,可在其叶片组织中形成碳酸钙晶体[7]。然而,目前关于不同无机碳来源在海草床无机碳库中的相对贡献仍缺乏定量评估,因此需要更多的案例研究,为海草床沉积物无机碳库的评估和管理提供数据支持。
与温带海域相比,热带和亚热带海域的海草床物种多样性更高,大型海草物种更为普遍,钙化生物的种类及数量也明显更多[8]。已有研究表明,以二药草属(Halodule)、泰来草属(Thalassia)和丝粉草属(Cymodocea)为优势种的热带或亚热带海草床,其沉积物中的无机碳储量通常较高[9]。位于海南省陵水县黎安港的海草床是我国首个省级海草床保护区[10]。作为我国热带混生型海草床的典型代表,该区域已发展成为海草床碳循环研究的热点区域。本研究以海南黎安港海草床为研究对象,旨在实现以下目标:1)量化不同海草物种分布区域及邻近裸地区域表层1 m沉积物的无机碳储量;2)利用碳酸盐碳、氧稳定同位素解析沉积物无机碳的主要来源及其贡献比例;3)结合已有的沉积物有机碳储量评估结果,探讨无机碳库对海草床整体碳汇评估的潜在影响。
1 材料与方法
1.1 研究区域及样品采集
黎安港位于海南省陵水黎族自治县黎安镇东南部(110.04°E—110.06°E,18.40°N—18.44°N),是一个天然的半封闭潟湖(图1)。该潟湖总面积约9.00 km2,平均水深较浅,大部分区域水深不足5 m,仅通过一条宽约60 m的潮汐汊道与外海相连,形成了相对独立的水体交换系统[11]。该地区属于热带海洋性季风气候,全年平均气温为25.4 ℃,降雨多集中在夏、秋两季。沉积物类型以砂为主,砂、粉砂和黏土的平均含量分别为73%、25%和2%[12]。潟湖内海草床总面积约为1.42 km2,主要分布在湖周0~3 m的浅水区域,由海菖蒲(Enhalus acoroides)、泰来草(Thalassia hemprichii)、圆叶丝粉草(Cymodocea rotundata)和卵叶喜盐草(Halophila ovalis)等海草构成[11]。其中,卵叶喜盐草在2019年的海草资源调查中仅在潟湖北部区域被发现[11],而在2021年的调查中,其在整个潟湖范围内均未被发现[13],故本研究未将与该物种相关的沉积物碳库纳入调查范围。
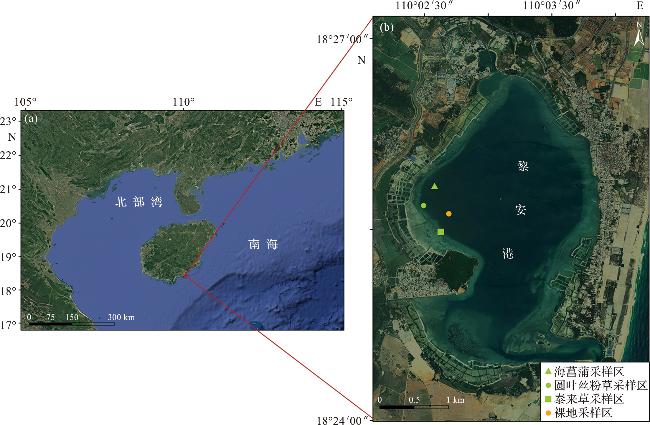
2023年6月—7月,分别在海菖蒲、泰来草、圆叶丝粉草分布区域及其邻近无植被裸地区域,各采集3根沉积物柱样,共计获得12根沉积物柱样(表1)。采样时,用橡胶锤将外径110 mm(内径103.6 mm)、长度700 mm的PVC管垂直敲击插入沉积物中,至无法继续推进为止,随后将沉积物柱样完整挖出,过程中尽量减少样品的压缩变形。现场对每根沉积物柱样以5 cm为间隔进行分样后分别装袋并编号。样品运回实验室后,在4 ℃条件下保存。
表1 黎安港沉积物柱样信息Tab.1 Information of sediment cores in Li’an Lagoon |
| 采集区域 | 柱样名称 | 柱样长度/cm | 分样数量/个 |
|---|---|---|---|
| 海菖蒲 分布区域 | LA-01 | 30 | 6 |
| LA-02 | 40 | 8 | |
| LA-03 | 40 | 8 | |
| 圆叶丝粉草 分布区域 | LA-04 | 45 | 9 |
| LA-05 | 50 | 10 | |
| LA-06 | 55 | 11 | |
| 泰来草 分布区域 | LA-07 | 60 | 12 |
| LA-08 | 60 | 12 | |
| LA-09 | 50 | 10 | |
| 裸地区域 | LA-10 | 55 | 11 |
| LA-11 | 50 | 10 | |
| LA-12 | 50 | 10 |
1.2 样品测定
取一定体积的沉积物样品,置于60 ℃烘箱中烘干至恒重,根据干重计算沉积物的容重。剔除干燥样品中的石块、残留植物根系等杂质,混匀、研磨并过筛。随后在室温条件下用足量2 mol·L-1盐酸溶液浸泡样品至无气泡产生,以完全去除样品中的CaCO3,之后用蒸馏水洗涤3次,再次烘干至恒重。使用元素分析仪(Thermo Scientific, FlashSmart)分别测定酸化处理前后沉积物样品中的碳含量,获得对应的总碳(TC)和有机碳(TOC)质量分数,两者差值即为沉积物样品中的无机碳(TIC)质量分数。计算公式如下:
ω(TIC)=ω(TC)-ωc(TOC)=ω(TC)-ωm(TOC)× $\left[1-\frac{100}{12}\omega \left(TIC\right)\right]$
整理后可得:
ω(TIC)= $\frac{3\left[\omega \left(TC\right)-{\omega }_{m}\left(TOC\right)\right]}{3-25{\omega }_{m}\left(TOC\right)}$
式中:ω(TIC)为无机碳的质量分数;ω(TC)为总碳的质量分数;ωc(TOC)为校正酸化质量损失后的有机碳质量分数;ωm(TOC)为元素分析仪实测的有机碳质量分数。
沉积物样品的无机碳或有机碳密度的计算公式如下:
ρ=ρd×ω
式中:ρ为沉积物样品的无机碳或有机碳密度,单位:g/cm3;ρd为各区域沉积物的平均容重,单位:g/cm3;ω为沉积物样品的无机碳或有机碳质量分数。
样品中碳酸钙的δ13C和δ18O值测定主要步骤如下:依据样品的无机碳含量称取约1 000~2 000 μg的研磨样品置于氦气氛围顶空瓶中,在70 ℃恒温条件下与无水磷酸反应40 min,以充分释放CO2气体,随后利用同位素比值质谱仪(Thermo Scientific Delta Q)分析所释放气体的碳、氧同位素比值。实验分析以国际原子能机构标准物质(IAEA-603)进行校准,测试精度为δ13C<0.01‰,δ18O<0.04‰。
1.3 数据分析
在呈现数据结果时,以单根沉积物柱样所有样品无机碳密度的平均值代表“柱样无机碳密度”,以各研究区域内3根柱样的无机碳密度平均值代表各区域的“区域无机碳密度”。将各区域的“区域无机碳密度”乘以1 m沉积厚度,并换算为国际单位(Mg·ha-1)后,即可获得对应研究区域表层1 m沉积物的无机碳储量(单位面积无机碳储量)。相关沉积物无机碳密度及无机碳储量结果均以平均值±标准差表示。数据处理与统计分析在 IBM SPSS Statistics 27.0.1软件中进行。首先检验数据是否符合正态分布与方差齐性要求;满足前提条件后,采用单因素方差分析(One-way ANOVA)分别对同一区域内部不同深度以及不同研究区域之间的无机碳密度进行显著性检验;若差异显著(即p<0.05),则采用Tukey HSD或Bonferroni方法对数据进行进一步的多重比较分析。
本研究中使用的同位素端元特征值参考自REN等[14]在同一区域测得的无机碳端元稳定碳、氧同位素值,具体包括:钙化底栖生物(δ13C:-0.21‰±0.19‰,δ18O:-2.79‰±0.07‰)、珊瑚礁(δ13C:-0.58‰±0.16‰,δ18O:-4.14‰±0.17‰)、钙化附生藻类(δ13C:-0.59‰±0.22‰,δ18O: -3.47‰±0.33‰)及陆源输入碳酸盐(δ13C: -9.43‰±1.63‰,δ18O:-10.91‰±0.91‰)。利用贝叶斯混合模型MixSIAR(基于R v.4.5.1)对研究区沉积物无机碳来源的贡献比例进行拟合与解析[15]。模型采用马尔可夫链蒙特卡罗(MCMC)算法进行复杂概率分布的近似采样。采用单因素方差分析检验三种海草分布区域及裸地区域之间不同无机碳来源贡献比例的差异,若p<0.05,则认为差异具有统计学显著性。
2 结果
2.1 沉积物无机碳含量与单位面积储量的差异
本研究中的三种海草分布区域及裸地区域的沉积物无机碳含量随深度的变化如图2所示。总体来看,相较于无海草覆盖的裸地区域,各海草分布区域的沉积物均表现出更高的无机碳含量(p<0.05)。其中,海菖蒲和圆叶丝粉草分布区域的无机碳含量显著高于泰来草分布区域(p<0.05)。具体而言,海菖蒲分布区域沉积物的无机碳含量最高,平均值为1.03%±0.10%(范围0.62%~1.30%);圆叶丝粉草分布区域沉积物的无机碳含量次之,平均值为0.82%±0.19%(范围0.26%~1.53%);泰来草分布区域沉积物的无机碳含量平均值为0.65%±0.13%(范围0.26%~1.08%);裸地区域的无机碳含量最低,平均值为0.60%±0.13%(范围0.23%~1.39%)。
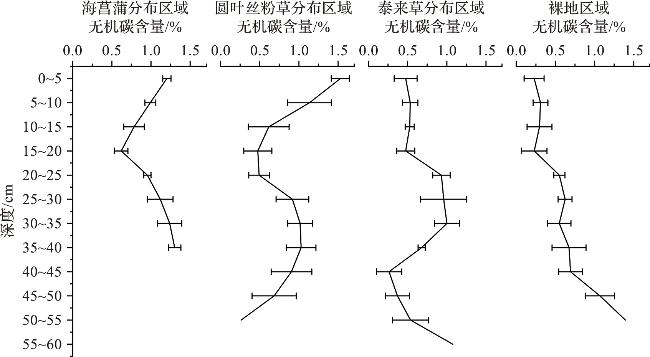
不同区域沉积物的无机碳含量随深度变化的趋势存在差异(图2)。具体而言,海菖蒲和圆叶丝粉草分布区域的变化趋势较为一致:无机碳含量在表层(0~20 cm)随深度增加而降低,在中层(20~40 cm)则转为上升;其中圆叶丝粉草分布区域的无机碳含量在深层(40~55 cm)再次随深度增加而下降。泰来草分布区域和裸地区域的无机碳含量在表层(0~20 cm)变化平缓;在中层(20~35 cm)随深度的增加无机碳含量略有上升;进入深层后(35~60 cm),两者变化趋势出现分异:泰来草分布区域的无机碳含量在35~45 cm深度范围内随深度的增加逐渐降低,在45~60 cm再次逐渐升高,而裸地区域的无机碳含量在35~60 cm深度内则随深度增加持续升高。
在同区域、不同深度间的沉积物无机碳密度比较中,圆叶丝粉草、泰来草分布区域及裸地区域不同深度沉积物的无机碳密度差异均未达到统计学显著水平(表2,p>0.05)。海菖蒲分布区域各深度沉积物的无机碳密度的差异在单因素方差分析中虽呈现显著性(p=0.034),但经Bonferroni法进行多重比较校正后,该显著性消失(p>0.1)。在不同区域间的沉积物无机碳储量比较中,各研究区域沉积物的无机碳密度差异显著,且呈现海草个体越大、对应沉积物无机碳储量越高的趋势(p<0.05)。具体表现为:海菖蒲分布区域的单位面积无机碳储量最高(105.66±9.45 Mg·ha-1),圆叶丝粉草分布区域次之(96.09±31.36 Mg·ha-1),二者分别比泰来草分布区域无机碳储量(75.05±8.17 Mg·ha-1)高出40.79%和28.03%;裸地区域的无机碳储量最低(59.84±24.55 Mg·ha-1),仅为有海草覆盖区域沉积物无机碳储量均值的64.85%。
表2 不同海草物种分布区域及裸地沉积物的无机碳密度与储量Tab.2 Inorganic carbon density and storage in sediments of different seagrass species distribution areas and bare areas |
| 采集区域 | 柱样名称 | 柱样无机碳密度a /(g·cm-3) | 区域无机碳密度b /(g·cm-3) | 表层沉积物单位面积无机 碳储量估算/(Mg·ha-1) |
|---|---|---|---|---|
| 海菖蒲分布区域 | LA-01 | 0.009 2±0.001 3 | 0.010 6±0.000 9 | 105.66±9.45 |
| LA-02 | 0.012 2±0.000 9 | |||
| LA-03 | 0.011 3±0.001 0 | |||
| 圆叶丝粉草分布区域 | LA-04 | 0.011 2±0.001 1 | 0.009 6±0.003 1 | 96.09±31.36 |
| LA-05 | 0.012 4±0.001 2 | |||
| LA-06 | 0.005 2±0.001 5 | |||
| 泰来草分布区域 | LA-07 | 0.007 0±0.001 0 | 0.007 5±0.000 8 | 75.05±8.17 |
| LA-08 | 0.0087±0.001 3 | |||
| LA-09 | 0.006 9±0.001 4 | |||
| 裸地区域 | LA-10 | 0.009 4±0.001 2 | 0.006 0±0.002 5 | 59.84±24.55 |
| LA-11 | 0.004 0±0.000 8 | |||
| LA-12 | 0.004 5±0.001 1 |
注:a.沉积物不同深度层间无机碳密度的显著性检验结果如下:海菖蒲分布区域差异显著(p=0.034;N=22),而圆叶丝粉草分布区域(p=0.173;N=30)、泰来草分布区域(p=0.079;N=33)及裸地区域(p=0.075;N=30)差异均未达到显著水平。b.不同区域之间(以柱样整体平均值为统计单元)沉积物无机碳密度差异均显著(p<0.05;N=12)。 |
2.2 沉积物碳酸盐碳、氧同位素组成特征
本研究中,海草床分布区域及其邻近裸地区域中沉积物柱样的碳、氧同位素值整体变化范围为δ13C:0.54‰~3.25‰,δ18O:-3.93‰~-2.63‰(图3)。海菖蒲分布区域、圆叶丝粉草分布区域、泰来草分布区域及裸地四个区域的δ13C和δ18O平均值分别为1.75‰±0.12‰和-3.34‰±0.07‰、1.70‰±0.10‰和-3.40‰±0.04‰、1.99‰±0.07‰和-3.38‰±0.04‰、1.64‰±0.09‰和-3.46‰±0.04‰。不同区域沉积物在垂向上(深度方向)的δ13C和δ18O值未呈现显著差异,其中海菖蒲分布区域沉积物的碳、氧同位素值变化幅度最大,裸地区域的同位素值变化幅度最小(图4)。
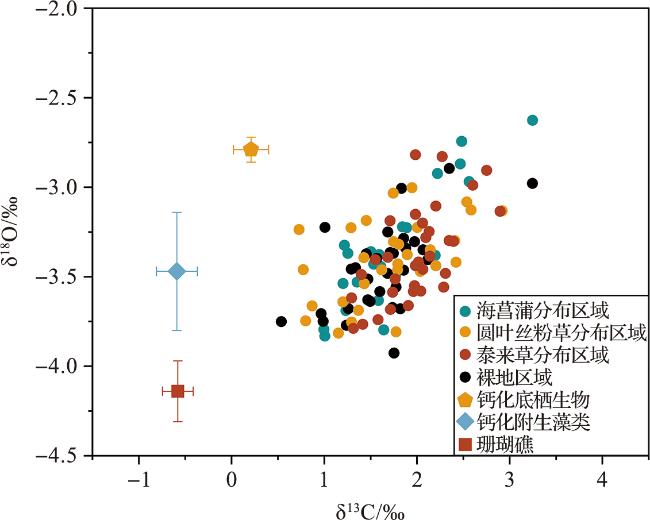
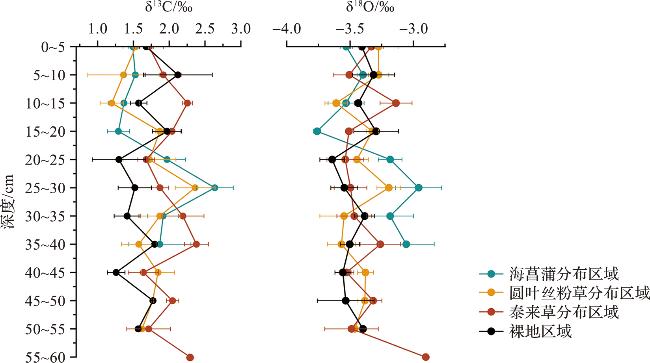
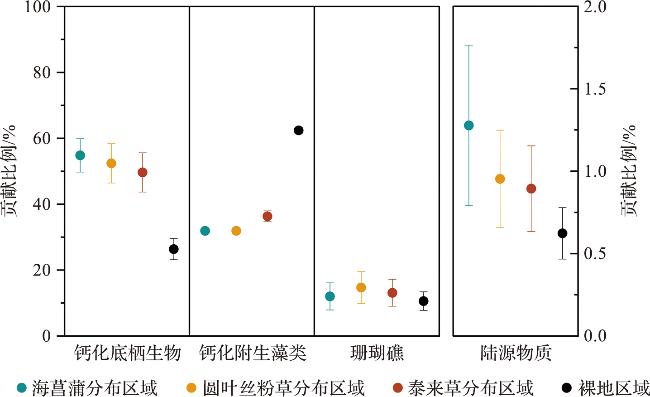
在此基础上,本研究参考REN等[14]在同一区域获得的钙化底栖生物、珊瑚礁、钙化附生藻类和陆源输入碳酸盐沉积物的δ13C和δ18O特征值,运用MixSIAR模型对研究区沉积物中碳酸盐的碳、氧同位素组成进行来源解析。结果表明,海草覆盖区域与裸地区域的无机碳来源具有显著差异(p<0.05)。海草覆盖区域,沉积物无机碳的贡献比例从高到低依次为:钙化底栖生物、钙化附生藻类、珊瑚礁和陆源物质,其平均贡献比例分别为49.66%~54.83%、31.90%~36.34%、12.00%~14.70%和0.89%~1.28%(图5)。而裸地区域沉积物无机碳的贡献比例从高到低依次为:钙化附生藻类、钙化底栖生物、珊瑚礁和陆源物质,其平均贡献比例分别为62.42%、26.37%、10.57%和0.62%。
3 讨论
3.1 海草物种对海草床沉积物无机碳积累的影响
不同海草物种在形态学上的差异是造成其碳捕获与封存能力分异的重要因素[16]。本研究结果显示(图2),海菖蒲和圆叶丝粉草分布区域表层沉积物的无机碳含量显著高于泰来草分布区域,这与海菖蒲和圆叶丝粉草个体显著大于泰来草的形态学特征相对应。较高的海草覆盖度和地上生物量可以通过阻滞水流促进颗粒物沉降,同时为钙化底栖生物提供栖息地、为钙化附生藻类提供附着基质,进而增强沉积物中无机碳的积累[11]。有研究表明,海菖蒲与圆叶丝粉草的平均根系长度(分别为15.15±2.08 cm与8.22±2.70 cm)明显长于泰来草(5.70±2.07 cm)[17]。较发达的地下茎-根系结构有助于增强沉积物稳定性,减少再悬浮,从而有利于已埋藏无机碳的保存[18]。相比之下,泰来草分布区域表层沉积物的无机碳含量较低,这很可能与其较稀疏的冠层结构、较短的地下茎-根系结构有关,这些形态特征限制了其捕获和促使颗粒物沉降的能力,导致部分无机碳沉积物在潮汐动力作用下发生流失。
3.2 沉积物无机碳积累的关键环境驱动因素:碳、氧同位素证据
基于MixSIAR模型的分析结果可知,以钙化底栖生物和钙化附生藻类为代表的内源无机碳是黎安港海草床沉积物中无机碳的主要来源,其平均贡献比例分别为49.66%~54.83%和31.90%~36.34%。其中,有海草分布区域的沉积物中以钙化底栖生物来源的无机碳为主,而邻近裸地区域沉积物中的无机碳则主要来源于钙化附生藻类。这一结果表明,海草的生长可以通过为生态系统中的钙化底栖生物和钙化附生藻类提供稳定的庇护环境和附着基底,直接促进钙化底栖生物和钙化附生藻类丰度的提升,从而增强生态系统中无机碳的原位生成与埋藏。相对而言,裸地沉积物中的无机碳则主要来自于邻近区域凋落的海草叶片及钙化附生藻类的异地埋藏[20]。
3.3 沉积物无机碳对海草床碳汇能力的影响
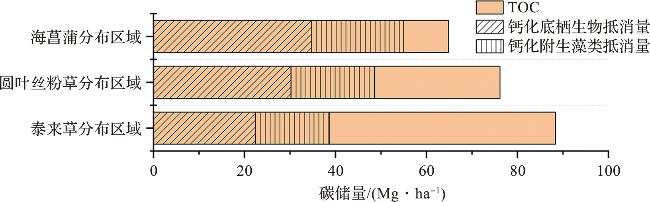
众多研究表明,海草床沉积物中含有丰富的无机碳。2021年对黎安港海草床有机碳储量的调查结果显示,其表层1 m沉积物的有机碳储量为69.61±26.11 Mg·ha-1 [13],与本研究的结果相近。而本研究结果显示,除泰来草分布区域外,海菖蒲与圆叶丝粉草分布区域内沉积物的无机碳储量均高于对应的有机碳储量,分别约为其1.63倍和1.26倍(表3)。潟湖中部的沉积柱样数据也表明,黎安港无机碳的埋藏速率比有机碳高出2.68倍[24]。在全球尺度上,海草床表层1 m沉积物的平均无机碳储量为654±24 Mg·ha-1,封存的无机碳总量约为11~39 Pg,相当于有机碳储量的5倍左右[9,25]。在富含碳酸钙的海草床(如地中海、佛罗里达湾和澳大利亚沿岸)中,无机碳含量显著高于有机碳的特征尤为突出[5,26-28]。
表3 不同区域与全球海草床有机碳/无机碳储量估算Tab.3 Estimation of organic/inorganic carbon stocks of seagrass meadows across different regions and globally |
| 区域 | 有机碳储量/ (Mg·ha-1) | 无机碳储量/ (Mg·ha-1) | 数据来源 |
|---|---|---|---|
| 巴西东南部 | 67.6±14.7 | 141.5±60.0 | 文献[5] |
| 佛罗里达湾(美国) | 175.0±20.4 | 754.6±26.7 | 文献[5] |
| 埃克苏马群岛 | 956±70.0 | 文献[29] | |
| 全球 | 120.8±3.5 | 654±24 | 文献 [9,25] |
| 海南黎安(海菖蒲) | 64.84±3.02 | 105.66±9.45 | 本研究 |
| 海南黎安(圆叶丝粉草) | 76.17±13.25 | 96.09±31.36 | 本研究 |
| 海南黎安(泰来草) | 88.37±4.91 | 75.05±8.17 | 本研究 |
注:所有数据均表征表层1 m沉积物储量。 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
需要指出的是,以上估算建立在“沉积物中钙化附生藻类和钙化底栖生物源的无机碳全部为原位形成”的简化假设之上,未考虑外源输入的影响。然而,在实际生态系统中,碳来源具有多源性,该假设可能低估了外源碳对沉积物碳库结构的贡献。例如,西班牙马略卡岛海草床中超过90%的无机碳为外源成分[3],我国南海近岸海草床的外源输入比例可达70%以上[14]。若低估外源有机碳的比例,则可能高估由碳酸钙原位生成引起的CO2释放对有机碳汇的抵消效应,从而低估海草床的实际净固碳能力。因此,准确量化沉积物无机碳中外源碳与内源碳的比例,也是评估海草床碳源-汇功能的关键。在未来的海草床固碳能力研究中,必须重视无机碳储量的准确测定与无机碳来源的精确解析。
4 小结
本研究以黎安港为例系统分析了典型混生型海草床沉积物的无机碳含量及其来源组成。总体而言,无机碳库在海草床碳收支中具有重要地位,对碳汇功能的准确评估具有关键意义。主要结论归纳如下。
1)黎安港海草分布区域的沉积物无机碳含量显著高于裸地区域,且呈现海草个体越大,对应沉积物无机碳含量越高的趋势(p<0.05),说明海草的生长有利于提高无机碳的捕集和埋藏效率。
2)该研究区海草床沉积物的无机碳主要来源于钙化底栖生物(平均贡献比例:49.66%~54.83%)和钙化附生藻类(31.90%~36.34%),而珊瑚碎屑(12.00%~14.70%)和陆源输送的外源无机碳(0.89%~1.28%)贡献较少。这表明在半封闭潟湖环境中,海草主要通过提高钙化生物(底栖生物与附生藻类)的丰度来提升沉积物无机碳的输入。
3)由于生态系统内原位碳酸钙(CaCO3)形成过程会伴随CO2释放,因此无机碳库的累积可部分抵消海草床的净碳汇强度。本研究的定量分析显示,黎安港海草床整体仍表现出显著的碳汇能力。在未来的研究中,准确量化沉积物无机碳储量、厘清沉积物无机碳中内源碳和外源碳的比例,是准确评估海草床碳汇功能的关键。