
0 引言
随着我国“双碳目标”的提出,对海岸带蓝碳生态系统碳汇能力的量化评估成为一项重要的工作。但是,在中国,海岸带蓝碳生态系统的碳汇强度和速度的计算方面,仍有待于进一步的数据支持[4]。近年来,我国在滨海湿地碳储量和沉积物碳埋藏的调查和数据分析方面取得了显著的进展[5⇓⇓-8],但相关参数的获得往往是基于长时间尺度(几十年到上百年)的平均态观测。从已有文献报道来看,我国红树林生态系统的固碳速率约为7~22万t/a;滨海盐沼生态系统的约为21~91万t/a;海草床生态系统的实测数据很少,有限数据表明固碳速率约为12万t/a[4,9-10]。在我国,滨海盐沼的总体固碳能力最强,主要是因为此类生态系统面积较大。值得注意的是,滨海盐沼的固碳能力估算值变化范围较大,这主要由两个原因引起:首先,滨海盐沼面积具有较大的不确定性,各项研究所采用的遥感影像识别结果存在差异,该问题有待于遥感解译标准的统一来解决;其次,短时间尺度,特别是年际尺度内碳汇能力观测,目前缺乏可靠的观测数据,需要进一步发展现场观测技术。
从碳汇能力的观测技术上来看,海岸带蓝碳的定量研究方法包括碳收支监测、模拟实验和模型研究[11],其中碳收支监测是实现碳核算的重要途径。海岸带蓝碳的固碳作用非常复杂,涉及到多个界面、多个尺度的过程。因此,如何定量化观测蓝碳系统的固碳能力,成为一个具有挑战性的问题。根据《巴厘岛行动计划》中对国家自主贡献清单需达到“可测量、可报告、可核查”的要求,对蓝碳生态系统开展现场观测是必需的工作要求。从已有的现场观测技术路线来看,基本上都是基于碳收支的原理,可以分为“碳通量”和“碳库”两大类[11]。“碳通量”观测是基于碳循环具体过程的观测,即观测各个过程的通量,最后得出总量,测量方法包括涡度法、箱法和稳定同位素法[11-12]。“碳库”观测则是将蓝碳生态系统假设为箱式模型,监测某一时间段内系统净固碳量,而不去考虑其中复杂的过程。这个方法相对简单,可以得出一年或几年的变化量,但无法度量各个过程对碳通量的贡献[11,13]。近年来,我国科学家在红树林和滨海盐沼湿地内布置了多个二氧化碳涡度塔进行碳通量的监测[11,14],但受限于观测设备以及后续数据处理、台站维护的高额成本,目前我国滨海湿地内的碳通量监测点十分有限。随着学科间交叉程度的深入,源于地貌学观测的方法和设备被引入到基于“碳库”法的碳收支监测中,用于量化观测沉积物碳埋藏速率[15-16]。其中的地表高程监测系统由固定桩和测量手臂组成,最早应用于地表沉降和相对海平面变化的研究,是目前测量地表高程变化最为精确的一种方法[16]。据此可以获得从季节到年际尺度的沉积速率,进而计算出沉积物碳埋藏速率,若结合植物参数的变化,可以获得滨海盐沼碳库的总体收支,完成对碳汇能力(本文以一定时间内的净碳汇量,即固碳速率为指标)的评估。
1 研究区域
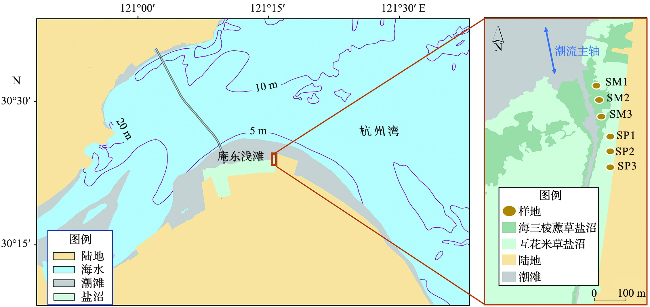
目前庵东浅滩滩面平均宽度达到6 km左右,上部分布有宽达3 km、以海三棱藨草和互花米草为优势种的盐沼。海三棱藨草是我国特有的具有代表性的盐沼先锋植物种属,是莎草科的一种多年生草本植物。互花米草是我国人工引种的潮滩植物,同样为多年生草本植物。庵东浅滩盐沼上部为互花米草优势区,盐沼的前缘发育了海三棱藨草区,在盐沼外部的潮间带则为淤涨型的淤泥质光滩[23]。
2 研究方法
综合考虑研究区域内植物分布特点、滩面高程变化的长期规律以及采样便利性等多个因素,选择庵东浅滩东部的典型盐沼区域作为布设固定样地的重点区域。沿着平行于潮流主轴的方向布设断面(如图1所示),断面穿越互花米草区和海三棱藨草区。在断面上布设固定样地6个,自海向陆,依次为海三棱藨草固定样地SM1、SM2、SM3和互花米草固定样地SP1、SP2、SP3。
本研究选择“碳库法”中的“碳库差异法”建立技术路线。“碳库差异法”是指调查起始时间和终止时间的调查区各碳库碳储量,两个时间点的总碳储量之差为调查区总碳汇量,该差值除以时间间隔和总面积即为单位面积内的固碳速率。该方法需要观测同一时间段内植物碳库和沉积物碳库的碳汇量变化,具体观测参数见表1。本研究的观测时间间隔为2个月,分别在2022年3月、5月、7月和9月开展了现场观测和样品采集。
表1 滨海盐沼碳库变化监测参数Tab.1 Measured parameters for coastal saltmarsh carbon stock changes |
| 监测内容 | 监测指标 | 监测方式 |
|---|---|---|
| 植物碳库增量 | 植物地上生物量 | 现场监测、现场采样、 室内分析 |
| 植物地下生物量 | ||
| 植物有机碳含量 | ||
| 沉积物碳库增量 | 地表高程 | 现场采样、室内分析 |
| 沉积物有机碳含量 | ||
| 沉积物容重 |
2.1 植物碳库增量监测
滨海盐沼主要被草本植物覆盖,草本植物地上部分在秋冬季节枯萎死亡。根据联合国政府间气候变化专门委员会(IPCC)关于滨海湿地温室气体排放清单的规定[29],盐沼植物地上部分的生物量变化不能计入到碳库增量变化中。因此,本研究中仅将地下生物量的增量计入碳库增量,地上部分的样方调查则用于分析植物的季节性变化。由于本文针对两种植物开展研究,对特定的植被碳库采用“植物碳库”的称呼。
在每个样地上设1个50 cm×50 cm的植物样方,现场开展盐沼植被种类组成、高度、数量调查,并齐地面割取样方内活体部分,带回实验室测定地上生物量。用采样器对样方内的沉积物进行采集,利用高压水枪分离出根、茎等生物活体(主要是根系)。根据已有研究,尽管植物根系可以到达1 m深度,但研究区盐沼的两种植物的主要根系分布在0~30 cm深度[25],因此本研究采集根系深度为30 cm。植物样品(茎、叶、根)带回实验室后用水冲洗多遍,剪碎后分别装入牛皮纸袋中,放入烘箱,于80 ℃烘干至恒重,测定干物质质量,从而计算单位面积植物地上部分和地下部分生物量。取适量烘干后的植物段(含茎和叶部分)、地下根系,分别用微型植物粉碎机粉碎,过100目标准筛;称取已过筛的植物样品,采用元素分析仪(德国Elementar Vario MICRO cube,测量绝对误差范围±0.30%)测定植物各组分的有机碳含量,获得碳转换系数。由于采集地下生物量对于固定样地的破坏较为明显,本研究中仅在3月和9月各测量一次地下生物量。
2.2 沉积物碳库增量监测
沉积物碳库增量主要用地表高程变化率(沉积速率)以及沉积物的容重、碳含量来共同计算。本研究采用美国Nolan地表高程监测系统进行沉积速率的观测,该设备精度可以达到±1.3 mm,超过现有的RTK-GPS的垂向高程测量精度,具体测量方法详见文献[16]。在地表高程监测点的周边,使用土壤取样器采集表层10 cm深的短柱状样,现场用样品袋包装好,带回实验室进行分样。一部分样品用于测定沉积物容重,另一部分沉积物样品装于干净玻璃或陶瓷器皿中,用Christ Delta 1-24 LSC型冷冻干燥机冻干并称重,之后使用元素分析仪(德国Elementar Vario MICRO cube)测定有机碳含量。
2.3 滨海盐沼碳库增量计算
碳库增量主要由固定样点内测得的两个时间节点(T1和T2)之间的碳库差值决定。植物碳库增量主要取决于地下生物量的年内变化,沉积物碳库增量取决于高程变化率、沉积物有机碳含量和容重的变化。单位面积内碳库增量具体计算公式如下:
CARV=ΔBioV×CV
CARS=10×SEC×BD×CS
CAR=10×SEC×BD×CS+ΔBioV×CV
式中:CARV为植物碳库增量,CARS为沉积物碳库增量,CAR为总碳库增量(单位均为g C·m-2·a-1)。ΔBioV为植物地下生物量增长率(单位:g·m-2·a-1),CV为地下各生物组分(主要是根系)的有机碳百分含量(单位:%),BD为表层(0~10 cm)沉积物的容重(单位:g·cm-3),CS为沉积物有机碳的百分含量(单位:%),SEC为地表高程变化率(单位:mm·a-1)。
3 结果
3.1 植物碳库的季节性变化
根据样方的调查结果(图2),海三棱藨草的植株高度、密度和生物量均具有显著的季节性变化。海三棱藨草从3月开始生长,3个站位的平均值显示,该季节植株高度仅达到2.7±1.5 cm,植株密度为210±43 株·m-2,地上生物量仅为0.5±0.1 g·m-2,地下生物量为17.1±10.7 g·m-2。海三棱藨草在5月和7月持续生长,并在7月底达到顶峰,植株高度达到51.0±2.7 cm,植株密度为1 656±338 株·m-2,地上生物量达到406.7±109.4 g·m-2。9月为生长季的结束期,植株高度、密度和地上生物量开始下降,出现明显的枯萎现象,但地下生物量达到高值,为72.9±72.2 g·m-2。海三棱藨草在观测期间没有随着生长发生自疏现象,地上生物量的增长依赖于植株高度和密度的增加,植物冠层具有高度低、密度高的特点。
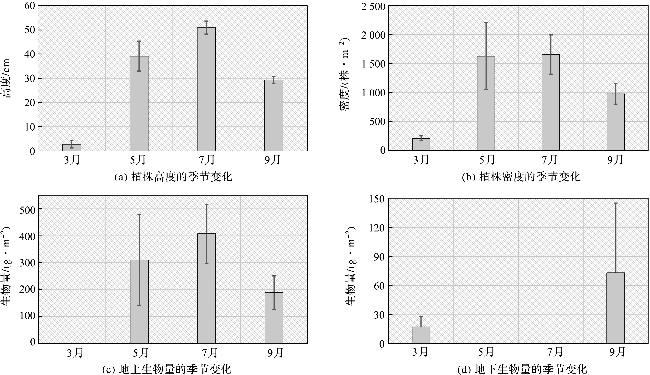
根据植物有机碳含量测量结果,海三棱藨草地上部分的有机碳平均含量为28%,地下部分的有机碳平均含量为19%。由计算可知,海三棱藨草生长顶峰期地上部分生物量碳可以达到114 g C·m-2,相当于418 g·m-2的CO2当量(转换系数为3.67),但是这部分有机碳随着植物的枯萎和凋落,通过分解和水动力运输,部分脱离了海三棱藨草蓝碳生态系统,返回到大气系统,部分分解后进入到沉积物碳库,另外一部分随着水动力输运,向外输出到其他生态系统中。因此从更长周期来看,就观测样地而言,地上部分固碳的贡献会间接进入到沉积物碳库部分,但本研究中不再计入。由于多年生植物的根茎持续生长,而且该部分地下生物量能持续累积,在植物生长季节内可以达到的生物量碳为11 g C·m-2,相当于40 g·m-2的CO2当量。
根据样方的调查结果(图3),互花米草的植株高度、密度和生物量均具有显著的季节性变化。互花米草从3月开始生长,3个站位的平均值显示,该季节植株高度仅达到40.3±15.4 cm,植株密度为120±39 株·m-2,地上生物量为383.3±72.5 g·m-2,地下生物量为119.0±51.3 g·m-2。互花米草在5月和7月持续生长,并在7月底达到顶峰,植株高度达到166.7±23.6 cm,但是植株密度从5月的195±42 株·m-2下降为7月的136±14 株·m-2,地上生物量在7月最高,达到3 607.9±2 030.1 g·m-2。9月为生长季的结束期,植株高度、密度和地上生物量开始下降,但是地下生物量达到高值,为1 516.5±305.4 g·m-2。值得注意的是,互花米草会随着生长和种内竞争发生自疏现象,地上生物量的增长依赖于植株高度和单株生物量的增加,和海三棱藨草相比,互花米草的冠层具有高度高、密度低的特点。
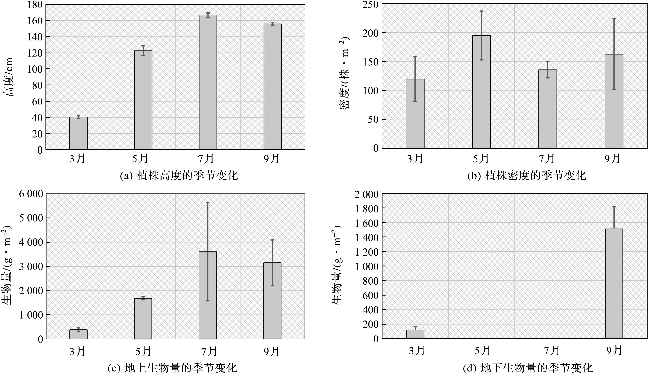
互花米草的地上部分有机碳平均含量为25%,地下部分为4%,说明地下部分的木质素含量较高。在生长顶峰期,互花米草地上部分固碳量可以达到902 g C·m-2,相当于3 310 g·m-2的CO2当量,但是这部分有机碳不计入碳库增量。在植物生长季节内,互花米草地下部分可以达到的固碳量为56 g C·m-2,相当于206 g·m-2的CO2当量。从地下生物量碳来看,互花米草的固碳能力强于海三棱藨草,主要原因是因为互花米草地下部分生物量增长较快,尽管其根系的有机碳含量较低。
3.2 沉积物碳库的季节性变化
根据固定监测点附近采集的沉积物样品的分析结果(图4a,4c)可知,海三棱藨草盐沼的沉积物容重和有机碳含量均具有比较明显的季节性变化。沉积物容重在3月和5月较高,在7月和9月较低。5月沉积物容重达到最高值,为1.41±0.14 g·cm-3;7月沉积物容重降至最低值,为1.06±0.03 g·cm-3。4次监测的平均容重为1.23 g·cm-3。这种容重的变化主要和季节性淹水时长变化及植物生长速度有关。海三棱藨草盐沼表层沉积物的有机碳含量也呈现较为显著的季节性变化。表层沉积物有机碳含量在3月和5月处于较低水平,为0.23%左右;在植物生长旺盛的7月和9月,表层沉积物的有机碳含量显著提高,7月最高,可达到(0.43±0.03)%,说明植物的生长对于沉积物碳库的增量具有重要贡献。4次监测获得的海三棱藨草盐沼表层沉积物平均有机碳含量为0.30%。
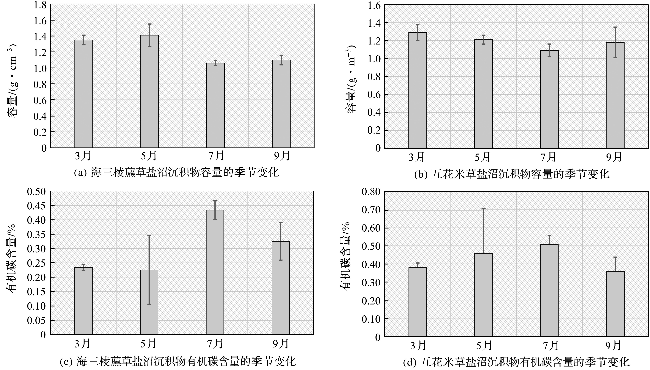
互花米草盐沼的沉积物容重和有机碳含量也具有比较明显的季节性变化(图4b,4d)。沉积物容重在3月和5月较高,在7月和9月较低。3月容重达到最高值,为1.29±0.09 g·cm-3;7月容重降至最低值,为1.09±0.07 g·cm-3。4次测量的平均容重为1.19 g·cm-3。产生这种变化的原因与海三棱藨草盐沼类似。互花米草盐沼表层沉积物的有机碳含量变化也有一定的季节性规律,但是不如海三棱藨草盐沼明显(图4c,4d)。表层沉积物有机碳含量在3月和9月处于相对较低的水平,最低值出现在9月,为(0.36±0.08)%;5月和7月,表层沉积物的有机碳含量有所上升,最高值出现在7月,可达到(0.51±0.05)%,说明植物的生长对于沉积物碳库的增量有一定的贡献。4次监测获得的互花米草盐沼表层沉积物平均有机碳含量为0.43%。相比海三棱藨草,互花米草的沉积物有机碳含量更高,而且在不同的季节相对稳定。
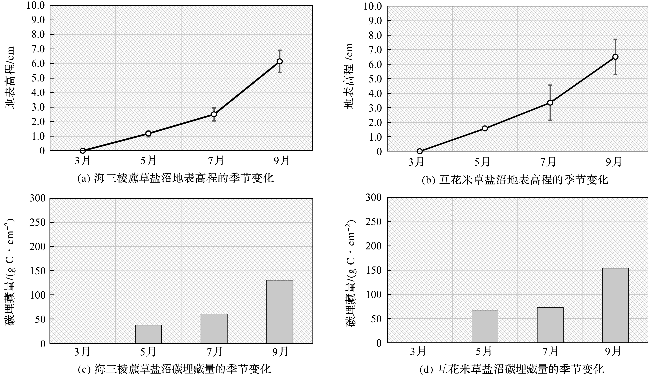
根据固定样地内地表高程监测系统的分析结果(图5),在监测期内,海三棱藨草和互花米草盐沼的地表高程均随着时间增加而增高,说明该区域为典型的淤积区域,有机碳在该区域发生净累积。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
3—9月,海三棱藨草盐沼各站位平均的沉积物表面高程变化达到6.15 cm。地表高程变化幅度在3—7月之间较小,为2.50 cm;在7—9月之间幅度较大,达到3.65 cm,这种差异与植物生长高峰期带来的有机质增加和沉积物捕获效应有关。由于监测时间为半年,可以计算得到海三棱藨草盐沼平均沉积速率为12.30 cm·a-1。
3—9月,互花米草盐沼各站位平均的沉积物表面高程变化达到6.51 cm。地表高程变化幅度在3—7月较小,合计为3.35 cm;在7—9月变幅较大,达到3.16 cm。互花米草盐沼平均沉积速率为13.02 cm·a-1,略高于海三棱藨草盐沼的沉积速率。
互花米草盐沼沉积物碳库的碳埋藏速率(588 g C·m-2·a-1),高于海三棱藨草盐沼(460 g C·m-2·a-1),主要原因是互花米草盐沼沉积物的有机碳含量显著高于海三棱藨草盐沼,同时沉积速率也略高于海三棱藨草盐沼。考虑到互花米草盐沼位于海三棱藨草盐沼的上部,外来有机碳颗粒的捕获受到海三棱藨草的阻挡,因此互花米草本身产生的有机碳向沉积物碳库转化的效率较高,这与前人对同一地区的沉积物有机碳来源分析结果一致[25]。
3.3 生态系统固碳能力
海三棱藨草和互花米草两种生态系统的净固碳能力是指植物固碳能力和沉积物碳埋藏能力之和,可以用固碳速率来表示这两类滨海盐沼生态系统的碳累积速度。由于海三棱藨草和互花米草都属于多年生草本植物,虽然碳库净增量不考虑地上生物量的增减[29],但是植物地下部分活体具有长期的固碳作用。因此,植物年际的固碳能力可以用地下生物固碳量来表示,而沉积物的碳埋藏速率则利用盐沼平均沉积速率外推至一整年,两者之和得到海三棱藨草和互花米草生态系统的固碳速率分别为471 g C·m-2·a-1和644 g C·m-2·a-1,海三棱藨草的固碳能力低于互花米草。事实上,此处使用的年际尺度固碳速率在植物部分比较合理,已经覆盖了植物的完整生长周期。相比之下,沉积物的碳埋藏速率则需要更长时间的测量(如2022年3月—2023年3月的测量数据)才可获得更为准确的数据。9月至次年3月期间草本植物凋落,尽管存在凋落物分解造成沉积物碳库在后续2个月内继续增长的可能性[30],但是植物地上部分死亡会造成潮滩捕获颗粒能力减弱[31],沉积物碳库整体的固碳能力应该有所减弱。因此本研究中的沉积物碳埋藏速率外推至1年时,存在估值过高的可能,未来需要开展更长时间尺度的观测加以补充。
4 讨论
滨海盐沼的固碳作用主要包括以下几个关键过程:1)植物通过光合作用固碳,这部分固碳作用可以用净初级生产力来表示;2)光合作用形成的有机碳向根系和沉积物碳库转移进行再分配,同时,植物的凋落物以及死亡后的残体有一部分残留在生态系统中,经腐殖化作用和泥炭化作用形成腐殖质和泥炭,也成为沉积物碳库的一部分,这部分碳累积具有一定滞后性;3)通过沉积动力过程形成的碳埋藏,即通过沉积物输运和沉降,使得有机碳在沉积物碳库中累积的过程,这个过程中还包括了外源有机碳的输入,特别是周边流域和海域的物质通过水平输运作用带来的有机碳输入。除了以上的碳汇过程以外,滨海湿地还存在碳排放的过程,包括植物呼吸作用、土壤呼吸作用、微生物分解、植物凋落物的分解、沉积物侵蚀作用、沉积物向外输运产生的输出、地下水循环带来的有机碳输出等[11-12,30,32⇓⇓-35]。由于本研究采用“碳库”法计算碳收支,因此暂不考虑水平输运带来的有机质交换,即不区分内源碳与外源碳。事实上,对于潮间带生态系统来说,水平输运带来的外源有机碳是非常重要的碳库增量来源。从该研究区域柱状样的碳氮比(C∶N)分析结果来看,该区域的沉积物碳库以海源的有机质输入为主[25]。如需进一步量化区分有机质的水平输运,则需要对沉积物中碳氮比值(C∶N)、稳定同位素组成(如δ13C)和生物标记物进行分析,通过端元模型进行外源输入的量化计算[30,34]。
植物的净初级生产力对于碳汇的贡献是前人研究的一个重点。邵学新 等[28]基于净初级生产力的现场观测,发现杭州湾南岸滨海湿地的海三棱藨草和互花米草植物群落的固碳能力分别达到274 g C·m-2·a-1和1 855 g C·m-2·a-1。类似的,王淑琼 等[36]发现崇明东滩的海三棱藨草和互花米草植物群落在生长季的固碳能力分别达到330 g C·m-2·a-1和1 320 g C·m-2·a-1。在本研究中,海三棱藨草地上生物量碳为114 g C·m-2,地下生物量碳为11 g C·m-2,相当于固碳能力达到125 g C·m-2·a-1。互花米草生长顶峰期地上生物量碳为902 g C·m-2,地下生物量碳为56 g C·m-2,相当于固碳能力达到958 g C·m-2·a-1。从整体上来看,本研究中的地上生物量碳观测值略低于前人的结果,主要归结为两个原因:首先,本研究没有计入凋落物部分的生物量;其次,本研究使用了实测植物有机碳含量来代替碳转换系数。
与此同时,多年生草本植物的地下部分始终存在活体部分,具有长期固碳的作用,应该予以考虑。前人研究发现,大多数滨海盐沼中的碳储存在地下活生物量和沉积物碳库中,这两个碳库通常被视为同一个碳库[37]。本研究试图将这两个碳库进行分离,分析各自的贡献。尽管部分植物根系经过分解以后进入到沉积物碳库已经被计算,但是活体部分(特别是活根),其有机碳含量远高于沉积物(约十倍至百倍),与沉积物碳库之间存在明显差异,若该部分不进行监测,则碳库增量会产生一定的低估。本研究尝试对两种典型盐沼植物的地下部分的固碳增量进行了观测与计算,发现虽然海三棱藨草和互花米草群落地下根系的碳库增量与沉积物碳库增量相比较小(两者之比分别为0.02和0.10),但这对整体碳库的固碳能力量化评估,特别是对地下生物量较高的植物群落(如芦苇和互花米草)来说,是一个有益的补充。这也说明若地下生物量变化不计入碳库监测,会给滨海盐沼碳库增量带来2%~10%的误差。
沉积物碳库是滨海湿地碳库的最主要组成,绝大多数的有机碳累积在沉积物碳库中[38]。生物地球化学和传统沉积学中的沉积物碳埋藏速率问题是“碳库”法的重要应用[39⇓-41]。植物碳库地上部分和地下部分通过植物死亡分解,均有一部分转移到沉积物碳库中,而且外源碳的输入也能在沉积物碳库中得到体现,因此沉积物碳埋藏速率是指示生态系统净固碳能力的一个重要指标。沉积物的碳埋藏速率取决于沉积速率、沉积物容重和有机碳含量,其中沉积速率是观测难度较大的一个指标。水平标志层法可以简便地测量滨海盐沼短期的沉积速率[16,42-43]。这个方法的缺点是不考虑压实作用带来的沉积速率变化,从而造成误差[16]。放射性元素210Pb和137Cs测年可用于滨海湿地几十年至百年时间尺度内的年代学测定和沉积速率测定[21,25,44],是目前用于长时间尺度沉积速率测定的主流方法。除此以外,近年来,地表高程监测系统在高精度的沉积速率观测方面发挥了重要的作用,其精度可以达到毫米级别[16]。考虑到地表高程监测系统中所采集的新淤积的沉积物含水量比较高,如果能用更高频的方法开展沉积通量监测来代替沉积速率,可更好地提高碳累积效率测定的准确性。值得注意的是,不同的沉积速率测定方法对应着不同的时间尺度,因此需要根据观测目的来确定合适的手段。
以杭州湾南岸滨海湿地为例,从柱状样210Pb测年分析发现该区域近几十年内的沉积速率为2.4~2.8 cm·a-1,结合沉积物测量结果,该地区海三棱藨草滩和互花米草滩的沉积物碳埋藏速率分别为140 g C·m-2·a-1和159 g C·m-2·a-1[25],远低于本研究中观测到的短期(年际)碳埋藏速率460 g C·m-2·a-1和588 g C·m-2·a-1。这主要是由于该地区的盐沼是近期(2010年之后)在庵东浅滩的潮滩中上部新发育的植物群落,原先为光滩状态[23],而放射性元素测年获得的是近几十年来的平均沉积速率,该沉积速率较短期的沉积速率偏低,最终导致碳埋藏速率在长期和短期尺度上存在较大的差异。在国家自主贡献清单的编制中,需要考虑到长期碳埋藏速率和短期碳埋藏速率之间的差异,并进行进一步结合,为制定技术路线提供较为明确的目标。另外,值得注意的是,不论是长期还是短期的碳埋藏速率观测,外来种互花米草的固碳能力均超过了本地种海三棱藨草,这对滨海湿地的保护和管理提出了新的挑战,即外来物种防控和海洋碳汇管理如何进行科学、有效的结合。
5 结论
本研究基于地表高程监测、现场调查与采样,结合实验室分析,对杭州湾南岸滨海盐沼内的本地种海三棱藨草群落和外来种互花米草群落开展了季节性观测,在分析植物碳库和沉积物碳库变化的基础上,量化评估了滨海湿地的固碳能力,取得了以下主要结论。
1)本地种海三棱藨草生长具有较强的季节性,地上植物部分生长季从3月开始,9月结束。在观测期内,地上部分固碳量达到114 g C·m-2,地下部分固碳量为11 g C·m-2。外来种互花米草的生长同样具有较强的季节性,地上部分固碳量达到902 g C·m-2,地下部分固碳量则为56 g C·m-2。
2)根据固定样地内地表高程监测系统的分析结果,在监测期内,海三棱藨草盐沼和互花米草盐沼的地表高程均随着时间增加而增高,说明该区域为典型的淤积区域,有机碳在该区域发生净累积。海三棱藨草盐沼平均沉积速率为12.30 cm·a-1,沉积物碳埋藏速率为460 g C·m-2·a-1。互花米草盐沼平均沉积速率为13.02 cm·a-1,沉积物碳埋藏速率为588 g C·m-2·a-1。两种植物群落的沉积速率和碳埋藏速率变化也具有一定的季节性,在夏、秋季达到高峰,这与植物生长有关。
3)对于多年生盐沼草本植物来说,尽管地上部分的净初级生产力固碳效率高,但是这部分有机碳从更长周期来看,没有固碳的贡献,而地下部分活体根系的固碳贡献则可以计入统计。将植物碳库和沉积物碳库的贡献相加,得到海三棱藨草盐沼和互花米草盐沼的固碳速率为471 g C·m-2·a-1和644 g C·m-2·a-1,表明海三棱藨草的固碳能力低于互花米草。
以上结论不仅提供了高精度的滨海盐沼碳库变化数据,为蓝碳的计量工作提供了支持,同时也从方法学和滨海湿地管理的角度提出了新的问题和挑战。不同的碳埋藏观测方法对应不同时间尺度和数据精度的碳汇量化观测需求,需要在管理目标的指引下去建立体系化的方法学。在滨海湿地保护和管理中,本地种和外来种固碳能力的差异从一个新的维度提出了新的管理问题。
致谢
自然资源部第二海洋研究所生态中心协助测量了有机碳的数据,洪秋鑫同学协助制作了部分图件,在此一并致谢!