
0 引言
为五种典型蓝碳生态系统,是推进生态系统固碳增汇的重点,也是建立健全蓝碳动态监测评估体系的重要内容①(① 浙江省自然资源厅.浙江省自然资源厅关于征求《浙江省自然资源领域海洋碳汇工作方案》意见的函. 2022-02-21.)。前人研究发现,澳大利亚东部沙岛的森林生态系统碳捕获能力高于相近位置的湿地生态系统和沼泽生态系统,辐射和温度对海岛森林生态系统的碳通量贡献度较高[3]。加拿大西北部湿地鱼岛在生长季节是净碳汇,光照水平和温度变化是昼夜净CO2吸收的控制因素,冰雪融化和物候是主要的季节性控制因素[4]。美国西南部和墨西哥之间的岛屿森林碳汇能力明显强于美国西部其他陆地森林[5]。海岛森林对净化空气、水土保持、涵养水源、调节气候等具有重要作用[6]。由于受地域限制、难以抵达、台风等因素影响,目前对海岛森林的生态监测,特别是碳汇功能监测十分缺乏,海岛森林尚未纳入蓝碳收支和森林碳汇估算体系中,可能是“未知碳汇”的重要组成部分[7-8]。因此,综合评估海岛森林碳循环过程的主要参量,包括净生态系统碳交换量(net ecosystem exchange, NEE)、总初级生产力(gross primary productivity, GPP)和生态系统呼吸(ecosystem respiration, Reco)将对补充海岛森林碳汇清单、建立我国蓝碳收支估算体系提供重要数据参考和理论支持。
本文以浙江省温州市平阳县南麂列岛国家级海洋自然保护区主岛南麂岛森林生态系统为研究对象,使用涡度相关技术定量评估该森林生态系统的碳汇功能,并探究南麂岛森林生态系统2020—2021年NEE、GPP与Reco的动态变化特征及其驱动因素,为建立我国蓝碳动态监测评估管理体系提供重要理论支撑。
1 区域概况与研究方法
1.1 研究区概况
南麂岛(27.476°N,121.053°E)是南麂列岛52个岛屿中最大的岛屿,面积为7.64 km2,海岸线长31.12 km[14,16],海湾众多,受海浪和潮汐侵蚀明显。南麂岛属典型的亚热带季风性湿润气候,四季分明,光照充足,雨量丰沛,空气湿润,年均降水量和气温分别为1 172 mm和16.5 ℃,极端气温为-4.5 ℃和34.3 ℃[14],降水主要集中在夏季。南麂岛属于基岩质海岛,土壤类型主要为红壤、粗骨土和海滨盐土,岩性主要为流纹质晶屑熔结凝灰岩[17]。南麂岛森林主要为常绿阔叶林,乔木优势树种主要为野梧桐、木麻黄、台湾相思等,灌丛优势树种主要有滨柃等,草本植物以菊科、莎草科、禾本科为主[17]。
1.2 数据来源
涡度相关塔(也称通量塔)站点设在南麂岛上,南麂岛森林面积约为4.85 km2[14],通量塔可监测的碳通量与水热通量有效距离为1 km。涡度相关测量系统由开放式的CO2/H2O红外气体分析仪(LI-7500A, LI-COR Lincoln, NE, USA)和三维超声风速仪(GILL-WM, Gill Instruments, UK)组成,分别测量CO2 、H2O的摩尔密度和风速、风向;采用Biomet系统(7900-101, LI-COR, Lincoln, NE, USA)监测环境变量[3,18]。通量塔高10 m,空气温度探头、湿度探头(HMP155, Vaisala, Helsinki, Finland)分别测量空气温度和相对湿度,安装高度为4 m,根据温度和相对湿度计算水气压差;雨量筒安装高度为10 m;土壤温度探头(7900-180, LI-COR, Lincoln, NE, USA)和土壤水分传感器(Stevens Hydra probe II, Stevens water monitoring systems, Oregon, USA)安装深度分别为地面以下0.05 m、0.1 m和0.2 m,本文采用的是0.05 m深处的土壤温度和水分数据;光合有效辐射由量子传感器(PPFD, LI-190r, LI-COR Bioscience, Lincoln, NE, USA)、四分量辐射计和光合辐射计监测,仪器安装高度为6 m[18]。数据采样频率为10 Hz,系统存储0.5 h碳、水通量的平均数据,由分析仪接口单元(LI-7550, LI-COR Biosciences, Lincoln, NE, USA)记录。由于雨量筒设备故障,降水数据观测结果偏差较大,为此收集了南麂岛气象台站(27.46°N,121.09°E)的湿度和累积降水数据。日均湿度、日累积降水量数据为南麂岛气象台站(http://data.cma.cn/)实时监测数据。涡度相关测量系统于2019年底开始工作,监测了2020—2021年南麂岛森林生态系统与大气间的碳通量与水热通量。

1.3 数据质量控制
涡度相关测量系统所测为10 Hz的高频数据,使用Eddypro软件进行计算和校正,处理成0.5 h的低频数据,主要包括二次坐标轴旋转[19]、趋势校正、空气密度校正[20]、超声波温度校正、统计检验、 频谱损失校正[21]等。受水汽、风速、天气变化和仪器设备故障等影响,Eddypro软件输出的0.5 h通量数据也会有一些“噪声”或是数据缺失,需要对数据进行质量控制,剔除“噪声”值,主要包括:1)剔除NEE质量标记为2的数据[22];2)剔除夜间NEE小于 0的数据、降雨量大于0的数据和CO2信号强度小于70的数据;3)排除超出合理阈值范围的异常值[23]。采用动点法确定摩擦风速u*阈值为0.61 m/s,去除夜间摩擦风速小于该阈值的通量数据[24]。为更好地处理异常值,计算7 d内有效观测数据的平均值(MEAN)和标准差(STD),若被检测值不属于(MEAN-3×STD,MEAN+3×STD)区间范围,则剔除。质量控制后最终保留了51.4%的有效NEE数据,与以往研究中有效数据量基本一致[25]。使用Reddyproc[9]软件包基于NEE有效数据的时间变化规律和总辐射(global radiation, Rg)、空气温度(air temperature, Tair)、饱和水汽压差(vapour pressure difference, VPD)3个环境因素对NEE数据进行插补。
1.4 数据分析
Reco通过下式进行计算:
Reco=rb×exp{E0[1/(Tref-T0)-1/(Tair-T0)]}
式中:Tref表示参考空气温度;Tair表示实际空气温度;T0表示初始空气温度,设为-46.02 ℃[27];rb表示参考温度下的基础呼吸(Tref为15 ℃);E0表示温度敏感性。
GPP基于Reco和NEE计算得到[26]:
GPP=Reco-NEE
NEE为负值表示吸收CO2,正值表示释放CO2。净生态系统生产力(net ecosystem productivity, NEP)与NEE的关系为:
NEP=-NEE
计算2020—2021年每日碳通量、环境因素的平均值,降水累积值,绘制碳通量和环境因子的变化趋势图。计算2020年和2021年各月份相同时刻NEE、GPP和Reco的平均值,得到不同季节各月份碳通量日变化,并按照春(3—5月)、夏(6—8月)、秋(9—11月)、冬(12—翌年2月)划分为四季,根据0.5 h碳通量累加得到年累积碳汇值。估算海岛森林日出、日落时间(基于Python程序的算法,名称为Solar.py)[28] 用于比较日尺度上CO2吸收开始和结束时主要的控制因素。基于主成分分析评估在0.5 h尺度上影响NEP、GPP与Reco的主要控制因素,每个变量对主成分轴的贡献可从载荷矩阵中得到[29]。为进一步确定环境因素与碳通量间的关系,基于2020—2021年GPP、Reco、NEP与环境因素的0.5 h有效数据,使用Excel 2013和Origin 2021软件计算不同环境因素和碳通量间的Pearson 相关系数并进行t检验,其中降水数据来源于气象站点日降水量的0.5 h平均值。本文数据制图皆基于软件Origin 2021。
2 结果与分析
2.1 海岛森林环境因子年际变化趋势
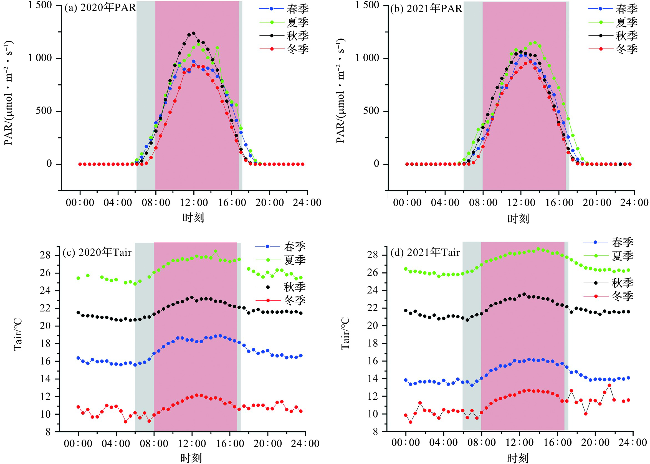
南麂岛属典型的亚热带季风性湿润气候,气象变量[空气温度(air temperature, Tair)、土壤温度(soil temperature, Tsoil)、降雨量(precipitation, P)、光合有效辐射(photosynthetically active radiation, PAR)、饱和水汽压差(vapour pressure difference, VPD)、相对湿度(relative humidity, RH)]和热通量[潜热(latent heat, LE)、显热(sensible heat, H)]都表现出明显的季节性(图2)。PAR的日均值变化范围为0~697.86 μmol·m-2·s-1,呈先上升后下降的单峰变化趋势,2020年和2021年PAR的最大日均值分别为697.86和648.43 μmol·m-2·s-1(图2)。Tair、Tsoil、P、VPD的日平均值变化趋势与PAR的日平均变化趋势较为一致(图2a~2c)。Tair和Tsoil的日均值变化范围分别为2.34~29.21 ℃和9.36~29.08 ℃;2020和2021年降水总量分别为1 042.5 mm和1 487.6 mm,两年的春季(3—5月)和夏季(6—8月)降水总量为1 626.3 mm,占两年降水总量的64.3%;VPD的最大日均值为5.64 kPa。RH在夏季较高,其他月份波动变化较大,其日均值在2020—2021年的变化范围为40%~100%(图2)。
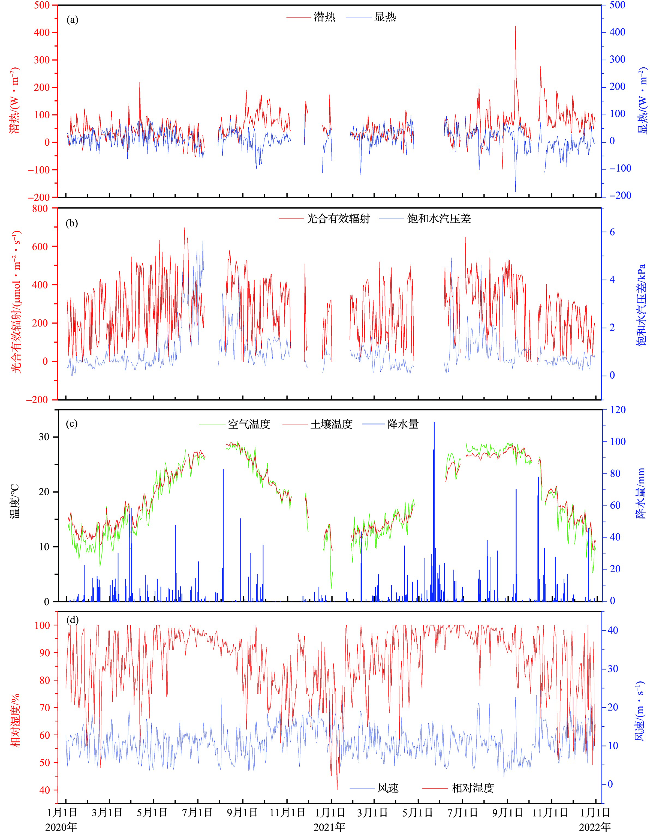
2.2 海岛森林NEE,GPP与Reco年际变化趋势
2020年和2021年海岛森林生态系统整体表现为“碳汇”,0.5 h NEE变化范围分别为-5.65~6.58 μmol·m-2·s-1和-6.99~8.66 μmol·m-2·s-1(图3a),年均NEE分别为-1.37 μmol·m-2·s-1和-1.58 μmol·m-2·s-1。2020年和2021年NEP分别为516 g C·m-2·a-1和598 g C·m-2·a-1,Reco分别为1 037 g C·m-2·a-1和1 646 g C·m-2·a-1,GPP分别为1 552 g C·m-2·a-1和2 244 g C·m-2·a-1(图3b~3c)。2021年4—9月0.5 h NEE数值相对2020年同期数值变化范围较大,且2021年累积碳通量变化速率也大于2020年;2021年GPP和Reco变化曲线出现多个峰值,但2020年仅在6月左右出现峰值(图3)。碳通量累积值变化规律明显,累积GPP和累积Reco逐日增加趋势基本一致,且GPP始终高于Reco,累积NEE始终为负值且逐日减小。2021年碳通量变化速率相对2020年更大,表明海岛森林累积碳汇功能逐渐增强。2020—2021年海岛森林累积NEP为1 114 g C·m-2,约占累积GPP(3 796 g C·m-2)的29.3%,即植被光合作用固碳量约有29.3%被存储下来。
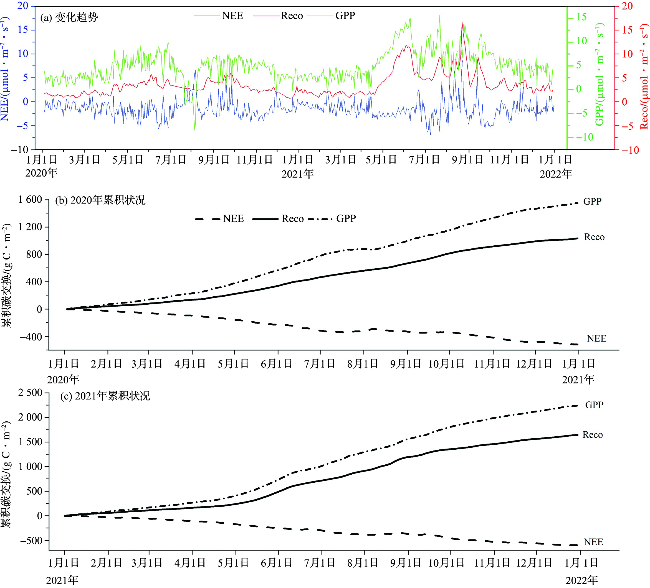
2.3 海岛森林NEE,GPP与Reco不同季节日变化趋势
2020—2021年南麂岛海岛森林总体发挥着“碳汇”功能,其中夏季和秋季碳汇作用较强,在日变化趋势图中NEE呈“U”型曲线,GPP和Reco呈倒“U”型曲线(图4)。2021年各季节的NEE最小值均低于2020年,各季节GPP和Reco日变化曲线整体高于2020年(图4)。NEE和GPP日变化幅度在冬季最小,秋季和夏季较大,说明秋季和夏季植被的光合作用能力较强(图4a~4d)。各季节NEE大约在8:00—17:00为负,即生态系统主要表现为吸收CO2,其他时间为正值即主要为释放CO2,且日间吸收CO2的强度明显大于夜间释放CO2的强度(图4a~4b)。日出后(约6:00),辐射和气温增加,光合作用逐渐增强,NEE由正值向负值转变,从8:00左右开始逐步实现CO2排放—净平衡—吸收的转变,在正午12:00左右NEE达到最小值,然后开始逐渐增大,并在日落前(约17:30)由负值转变为正值,开始释放CO2。夏季和秋季海岛森林生态系统日间CO2吸收通量和夜间CO2释放通量相对较大,出现了不同的吸收峰值。
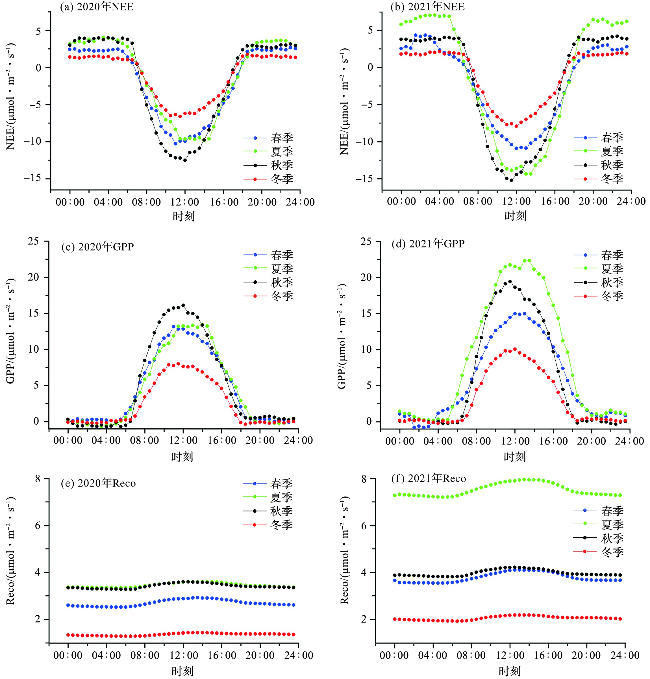
GPP的日变化呈抛物线,中午11:00—13:00,海岛森林光合作用强度达到峰值,夏季和秋季高于春季和冬季。2020年夏季和秋季Reco较为一致,而秋季的GPP较高,因此海岛森林在秋季的碳汇功能强于其他季节。2021年海岛森林夏季Reco显著高于其他季节,而秋季较低的Reco和较高的GPP促使海岛森林在秋季发挥较强的碳汇功能(NEE最小值为-15.23 μmol·m-2·s-1),与夏季(NEE最小值为-14.35 μmol·m-2·s-1)几乎相当。
海岛森林日出至日落时间为6:00—17:30,碳吸收(NEE<0)时间为8:00—17:00,相对光合作用发生时间,海岛森林发挥碳汇功能的时间滞后。结合PAR和Tair的日变化趋势(图5)分析可知,日出前和日落后Tair相对较低,太阳辐射较弱,PAR和光合作用强度较弱,生态系统呼吸作用大于光合作用,海岛森林表现为释放CO2;当Tair达到10.05~27.76 ℃,PAR达到110.47~429.44 μmol·m-2·s-1时,海岛森林光合作用强度高于生态系统呼吸作用,表现为吸收CO2。日落时Tair相对日出时更高,植物气孔导度较小,呼吸作用较强[30],因此,日落时从CO2吸收转变为CO2释放经历的时间较短。
2.4 2020—2021年海岛森林碳通量的影响因素
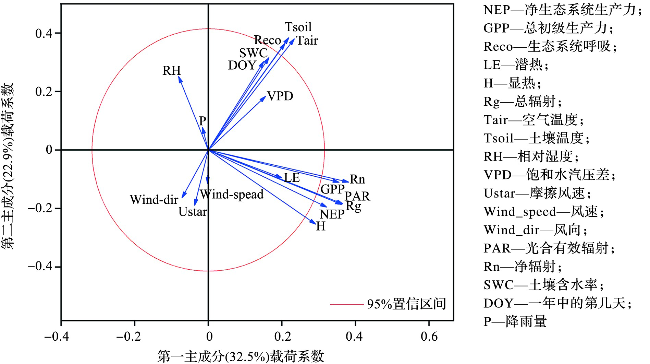
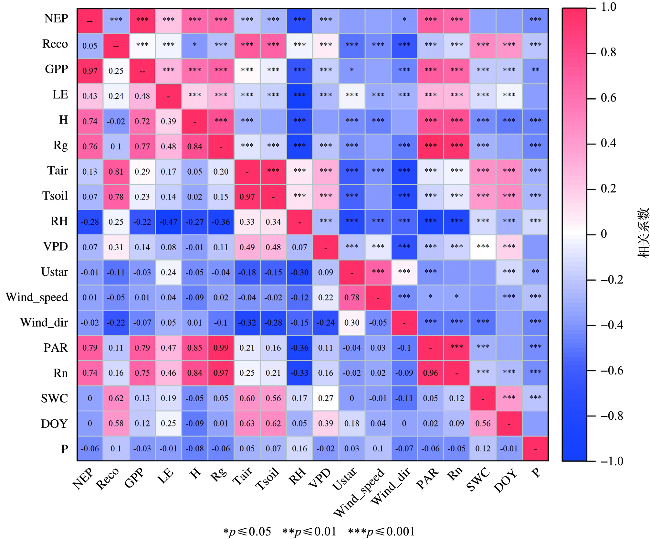
各环境因子与NEP、GPP和Reco的相关性结果显示(图7),NEP与GPP显著正相关,相关系数为0.97(p≤0.001),表明呼吸强度相同的情况下,NEP随GPP的增强而增加,即随光合作用的增强而增强。GPP与H、Rg、PAR、Rn皆呈显著正相关,相关系数变化范围为0.72~0.79(p≤0.001),即这4个因子会影响GPP进而影响NEP的大小。Reco与Tair、Tsoil、SWC、DOY(表示一年中的第几天,day of year)显著正相关(p≤0.001),相关系数变化范围为0.58~0.81,表明水热因素(特别是温度)对南麂岛森林碳储量有重要影响,这与前人研究结果一致[25,31]。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
3 讨论
3.1 海岛森林生态系统碳汇功能分析
受降水量和气温等环境因素影响,南麂岛2021年NEP(598 g C·m-2·a-1)高于2020年(516 g C·m-2·a-1),两年平均值为557 g C·m-2·a-1,与以往海岛森林研究结果(359~651 g C·m-2·a-1)较为一致(表1)。受气候环境、水文条件和地理位置等综合因素影响,红树林、盐沼湿地和海草床的NEP表现出较大的地域性差异,其NEP变化范围分别为73.8~2 101.77 g C·m-2·a-1、237~901 g C·m-2·a-1和367~646 g C·m-2·a-1(表1)。统计分析表明,全球范围内红树林、盐沼、海草床三类海岸带蓝碳生态系统和海岛森林生态系统的碳汇通量中值分别为721 g C·m-2·a-1、522 g C·m-2·a-1、507 g C·m-2·a-1和503 g C·m-2·a-1,南麂岛海岛森林年均碳汇通量分别为红树林、盐沼、海草床和海岛森林碳汇通量的77%、107%、110%和111%,表明南麂岛海岛森林年均碳汇通量相对其他3类海岸带蓝碳生态系统处于较高水平,提高海岛森林固碳潜力评估的精确度将有利于完善我国海洋蓝碳收支估算体系。
表1 全球范围内典型地区海岸带蓝碳生态系统的净生态系统生产力 (NEP)Tab.1 NEP in blue carbon ecosystems of coastal zone in typical regions on a global scale |
| 研究区域 | 经纬度 | 蓝碳类型 | NEP/(g C·m-2·a-1) | 参考文献 |
|---|---|---|---|---|
| 广东高桥 | 21.56°N, 109.75°E | 自然红树林 | 2 101.77 | 文献[32] |
| 福建云霄 | 23.92°N, 117.41°E | 自然红树林 | 540~1 233 | 文献[25] |
| 广东雷州 | 20.00°N, 110.09°E | 恢复红树林 | 1 105.15 | 文献[32] |
| 香港米埔 | 22.49°N, 114.3°E | 恢复红树林 | 690~890 | 文献[33] |
| 广东高桥 | 21.56°N, 109.75°E | 自然红树林 | 721 | 文献[34] |
| 广东珠江口 | 22.60°N, 113.64°E | 恢复红树林 | 175 | 文献[35] |
| 中国 | 红树林 | 209~973 | 文献[10,13] | |
| Florida Everglades, USA | 25.364 6°N, 81.077 9°W | 自然红树林 | 832~1 170 | 文献[36] |
| Sundarbans, India | 20.82°N, 88.61°E | 自然红树林 | 249 | 文献[37] |
| New Caledonia, France | 20.937 5°N, 164.658 3°E | 自然红树林 | 73.8 | 文献[38] |
| 全球 | 红树林 | 649.89 | 文献[39] | |
| 上海长江口 | 31.213 0°N, 121.906 9°E | 盐沼湿地 | 901 | 文献[40] |
| 上海九段沙 | 31.213 0°N, 121.906 9°E | 盐沼湿地 | 550~869 | 文献[41] |
| 上海崇明东滩 | 31.516 6°—31.516 9°N, 121.960 0°—121.971 6°E | 盐沼湿地 | 518~737 | 文献[42] |
| 山东黄河三角洲 | 37.763 8°N, 118.973 3°E | 盐沼湿地 | 237~261 | 文献[43] |
| 中国 | 盐沼湿地 | 865 | 文献[12] | |
| Massachusetts, USA | 42.736°N, 70.828°W | 盐沼湿地 | 256~337 | 文献[18] |
| 全球 | 盐沼 | 522.17 | 文献[39] | |
| 全球 | 海草床 | 367~646 | 文献[11] | |
| Tucson, USA Arizona, USA | 32.416 6°N, 110.725 3°W | 海岛森林 | 371~651 | 文献[5] |
| Vancouver Island, Canada | 49.519 4°—49.872 2°N, 124.901 6°—125.335 0°W | 海岛森林 | 359 ~635 | 文献[44] |
| 浙江南麂岛 | 27.476°N, 121.053°E | 海岛森林 | 516~598 | 本研究 |
注:表中自然红树林是指野生红树林,恢复红树林是指人为参与种植或培育的红树林。以往研究在统计全球或中国的NEP时,未指明红树林、盐沼或海草床的类型,故此表格中也未作区分。 |
3.2 海岛森林生态系统碳通量影响因素分析
南麂岛海岛森林碳汇通量受总辐射(Rg)、光合有效辐射(PAR)、净辐射(Rn)、显热(H)、土壤温度(Tsoil)和空气温度(Tair)等综合因素的影响。H、Rg、PAR、Rn与NEP和GPP呈显著正相关(p≤0.001)(图6~图7)。太阳辐射是植被光合作用的必要条件,且森林的冠层结构复杂,太阳辐射增加会让更多的叶片暴露在漫反射的辐射光线下,参与光合作用的树冠也更多,进而使得GPP升高[45]。南麂岛森林主要为常绿阔叶林,植被覆盖的季节性变化较小,太阳辐射会促进植被吸收CO2。PAR表现为NEP的主要驱动因素,在不同生态系统中,NEP对PAR的响应趋势受特定因素(如VPD和Tair)的阈值限定[32,38],在本研究中NEP与VPD和Tair不相关(p≤0.001)(图7)。0.5 h尺度上H与GPP显著正相关,H是评价植被光合作用过程中水分胁迫的重要指标[46]。南麂岛森林的温度变化显著影响Reco,Tair和Tsoil与Reco呈显著正相关,与GPP的相关性较弱(图7)。水分增加可能会限制土壤中气体或物质的扩散速率[26,47],也可能促进碳吸收[39]。0.5 h尺度上南麂岛森林SWC对NEP的影响并不显著,这是因为研究区紧邻海洋,短时间内水分条件变化对NEP的影响相对辐射因素和温度因素较弱[48]。
4 结论与展望
本文基于涡度相关数据探究了2020—2021年南麂岛森林生态系统的固碳速率,揭示了CO2通量与环境因子的关系,得出如下结论。
1)南麂岛森林生态系统表现为“碳汇”,2020年和2021年NEP分别为516 g C·m-2·a-1和598 g C·m-2·a-1,Reco分别为1 037 g C·m-2·a-1和1 646 g C·m-2·a-1,GPP分别为1 552 g C·m-2·a-1和2 244 g C·m-2·a-1。
2)海岛森林日出至日落的时间为6:00—17:30,碳吸收的时间为8:00—17:00。日出、日落时较低的Tair和太阳辐射,是导致光合作用时间和碳吸收时间不一致的主要原因。
3)主成分分析结果表明,2020—2021年太阳总辐射(Rg)、光合有效辐射(PAR)、净辐射(Rn)、显热(H)与NEP和GPP呈显著正相关(p≤0.001),促进CO2吸收;空气温度(Tair)和土壤温度(Tsoil)与Reco呈显著正相关(p≤0.001),促进CO2排放。
海岛是维护国家海洋权益、保护海洋生态的桥头堡,海岛森林发挥着净化空气、水土保持、涵养水源、调节气候等重要作用,尤其对海岛保护与可持续发展意义重大。由于受地域限制、难以抵达、台风等影响,海岛森林碳汇储量尚未纳入岛群生态系统碳储量和碳汇能力估算体系中。基于涡度相关技术监测海岛森林碳通量将为海岛生态系统碳汇评估提供重要的数据基础与理论支持,可为浙江省自然资源领域海洋碳汇工作方案落实提供重要参考。