
0 引言
赤杆菌科(Erythrobacteraceae)细菌隶属于α-变形杆菌纲(Alphaproteobacteria)、鞘氨醇单胞菌目(Sphingomonadales),包含了19个具有有效发表名称的属(https://lpsn.dsmz.de/family/erythrobacteraceae)。赤杆菌科菌株广泛分布于海洋环境,目前已在海水、海洋蓝藻垫、潮滩、红树林沉积物、热泉、深海沉积物和海洋无脊椎动物等不同生境中分离得到该科菌株[1⇓-3]。因其营好氧异养生长,且具光合作用功能,被视为好氧不产氧光合异养菌(aerobic anoxygenic phototrophic bacteria, AAPB),在全球海洋碳循环和能量流动过程中发挥着重要作用。赤杆菌科菌株可合成多种类胡萝卜素,使其呈红色、橙色、黄色与棕色等颜色[3-4],且海洋中不同生境来源的赤杆菌科细菌的类胡萝卜素合成基因种类及结构存在着一定的差异[5],这可能与它们在不同海洋环境中的生存策略有关。深海环境具有无光照、静水压高、温度低和含氧量高等特征,类胡萝卜素有助于微生物适应这一极端环境[6]。MATSUMOTO等[7]在日本冲绳石垣岛海岸沉积物中发现嗜盐菌Altererythrobacter ishigakiensis JPCCMB0017可产生具有抗氧化功能的虾青素。NIERO等[8]从西南大西洋深层海水中分离获得赤杆菌科菌株Erythrobacter citreus LAMA 915,发现该菌会积累类胡萝卜素,以应对强氧化胁迫,提高了菌株在深海环境的竞争优势。因此,研究海洋赤杆菌科细菌的类胡萝卜素合成机制对了解赤杆菌的生长和代谢机制,并将其应用于生物技术领域具有重要的科学意义。
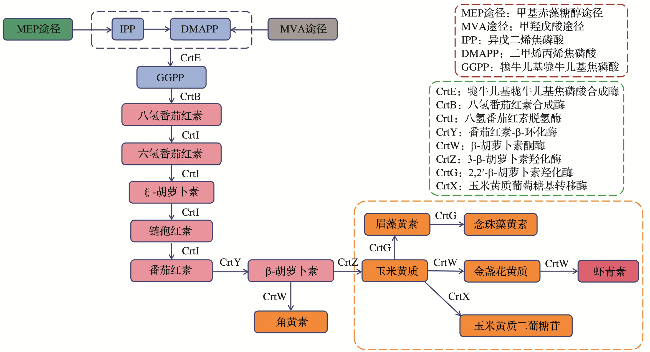
目前类胡萝卜素在细菌中的生物合成途径主要为两种(图1)[9-10],分别为通过甲羟戊酸途径(mevalonate pathway,MVA 途径)和甲基赤藻糖醇途径(2-C-methyl-D-erythritol 4-phosphate pathway,MEP 途径)合成所有已知类胡萝卜素的共同前体物——异戊二烯焦磷酸(isopentenyl pyrophosphate,IPP)及其异构体二甲烯丙烯焦磷酸(dimethylallyl pyrophosphate,DMAPP)[11]。随后再经过牻牛儿基牻牛儿基焦磷酸合成酶基因(crtE)、八氢番茄红素合成酶基因(crtB)、八氢番茄红素脱氢酶基因(crtI)、番茄红素-β-环化酶基因(crtY)、3-β-胡萝卜素羟化酶基因(crtZ)、β-胡萝卜素酮酶基因(crtW)、2,2'-β-胡萝卜素羟化酶基因(crtG)、玉米黄质葡萄糖基转移酶基因(crtX)等一系列与类胡萝卜素合成相关基因的编码蛋白催化形成不同类型的类胡萝卜素[12]。近年来,随着基因组技术的快速发展,大量海洋赤杆菌科菌株基因组已获测序,因此可开展基因组水平上的类胡萝卜素合成机制研究。开展海洋赤杆菌科细菌的类胡萝卜素合成基因多样性和进化模型研究,有助于评估该功能基因的基因多样性和进化机制,然而目前尚未有研究对海洋赤杆菌科细菌的类胡萝卜素合成基因进行系统分析。本研究选取了107株浅海和深海来源的赤杆菌科菌株,探究类胡萝卜素合成基因(crtBEGIWXYZ)在赤杆菌科中的分布情况,并根据类胡萝卜素合成基因的氨基酸序列开展系统发育分析,剖析浅海和深海来源赤杆菌科菌株的类胡萝卜素合成基因的存在和分布差异,解析海洋赤杆菌科菌株中类胡萝卜素的合成在不同环境来源下的适应进化机制,有助于全面认识海洋中产类胡萝卜素的菌株资源及其功能基因的分布与进化特征。
1 材料与方法
1.1 赤杆菌科基因组获取及基因组系统发育分析
基于基因组分类数据库(Genome Taxonomy Data-base, GTDB, https://gtdb.ecogenomic.org/)和美国国家生物技术信息中心(National Center for Biotechnology Information, NCBI, https://www.ncbi.nlm.nih.gov/)数据,获取107株赤杆菌科菌株及系外群(Rhodospirillum rubrum ATCC 11170T)的基因组,菌株基因组信息如表1所示。采用FastANI v1.0软件[13]计算基因组平均核苷酸一致性(average nucleotide identity, ANI),比较基因组相关性。使用Prokka v.1.14.5软件[14],对基因组注释并获得编码序列(coding sequences,CDSs)。基因组分析方法参照前期研究[15],使用Proteinortho v.5.16b软件[16]分析直系同源簇(orthologous clusters, OCs),参数为“-e 1e-5 -cov=50 -identity=50”,以获取单拷贝OCs。
表1 107株赤杆菌科菌株基因组信息Tab.1 Genomic information for 107 Erythrobacteraceae strains in this study |
| 序号 | GenBank登录号 | 菌株 | 基因组大小/bp | 基因组GC含量/% | Contig数量/个 | 来源 |
|---|---|---|---|---|---|---|
| 1 | GCA_001626395.1 | Erythrobacter sp. REDSEA-S40_B1 | 2 688 120 | 63.7 | 7 | 浅海海水 |
| 2 | GCA_001626415.1 | Erythrobacter sp. REDSEA-S28_B2 | 2 687 817 | 63.7 | 8 | 浅海海水 |
| 3 | GCA_001626515.1 | Erythrobacter sp. REDSEA-S43_B2 | 2 871 770 | 63.6 | 18 | 浅海海水 |
| 4 | GCA_003149575.1 | Altererythrobacter sp. XM-24bin4 | 2 534 571 | 56.8 | 12 | 浅海海水 |
| 5 | GCA_014983025.1 | Erythrobacter sp. E2-1 Yellow Sea | 2 518 489 | 57.4 | 31 | 浅海海水 |
| 6 | GCA_017644245.1 | Altererythrobacter sp. HCUG1430-4 | 2 509 776 | 56.8 | 9 | 浅海海水 |
| 7 | GCF_000152865.1 | Erythrobacter sp. NAP1 | 3 266 173 | 61.0 | 2 | 浅海海水 |
| 8 | GCF_000176355.1 | Alteriqipengyuania bathyomarina JL354 | 3 273 334 | 65.0 | 68 | 浅海海水 |
| 9 | GCF_000186705.1 | Alteriqipengyuania sp. JLT1363 | 3 117 324 | 64.9 | 26 | 浅海海水 |
| 10 | GCF_000235975.1 | Novosphingobium pentaromativorans US6-1 | 5 344 974 | 63.1 | 123 | 浅海沉积物 |
| 11 | GCF_000767465.1 | Novosphingobium pentaromativorans US6-1 | 5 457 578 | 63.0 | 6 | 浅海沉积物 |
| 12 | GCF_000813185.1 | Novosphingobium sp. MBES04 | 5 361 448 | 63.3 | 33 | 浅海海水 |
| 13 | GCF_001008165.2 | Croceibacterium atlanticum 26DY36 | 3 475 106 | 61.9 | 2 | 深海沉积物 |
| 14 | GCF_001010925.1 | Aurantiacibacter gangjinensis K7-2 | 2 724 959 | 62.7 | 8 | 浅海海水 |
| 15 | GCF_001010945.1 | Alteraurantiacibacter luteus KA37 | 2 887 216 | 67.2 | 22 | 浅海沉积物 |
| 16 | GCF_001013305.1 | Alteraurantiacibacter marinus HWDM-33 | 2 834 102 | 59.1 | 10 | 浅海海水 |
| 17 | GCF_001021555.1 | Alteraurantiacibacter marinus KCTC 23554 | 2 835 898 | 59.1 | 5 | 浅海海水 |
| 18 | GCF_001028625.1 | Pelagerythrobacter marensis KCTC 22370 | 2 885 033 | 64.7 | 1 | 浅海海水 |
| 19 | GCF_001077815.2 | Aurantiacibacter atlanticus s21-N3 | 3 226 359 | 58.3 | 2 | 深海沉积物 |
| 20 | GCF_001281485.1 | Altererythrobacter epoxidivorans CGMCC 1.7731 | 2 786 256 | 61.5 | 1 | 深海沉积物 |
| 21 | GCF_001461885.1 | Pelagerythrobacter marensis DSM 21428 | 2 902 055 | 64.7 | 14 | 浅海海水 |
| 22 | GCF_001634625.1 | Croceicoccus bisphenolivorans H4 | 3 602 538 | 62.8 | 74 | 浅海海水 |
| 23 | GCF_001634715.1 | Erythrobacter sp. HI0020 | 3 189 298 | 63.5 | 275 | 浅海海水 |
| 24 | GCF_001635465.1 | Erythrobacter sp. HI00D59 | 3 347 358 | 63.4 | 121 | 深海海水 |
| 25 | GCF_001635685.1 | Erythrobacter sp. HI0063 | 3 258 146 | 63.3 | 356 | 深海海水 |
| 26 | GCF_001661675.2 | Croceicoccus marinus E4A9 | 4 109 188 | 64.5 | 3 | 深海沉积物 |
| 27 | GCF_001661915.1 | Croceicoccus pelagius Ery9 | 3 306 530 | 62.8 | 40 | 浅海海水 |
| 28 | GCF_001661965.1 | Croceicoccus mobilis Ery22 | 4 210 079 | 62.5 | 32 | 深海沉积物 |
| 29 | GCF_001663155.1 | Altererythrobacter ishigakiensis NBRC 107699 | 2 673 978 | 56.9 | 1 | 浅海沉积物 |
| 30 | GCF_001677335.1 | Erythrobacter dokdonensis DSW-74 | 2 995 154 | 64.8 | 11 | 浅海海水 |
| 31 | GCF_001687545.1 | Paraurantiacibacter namhicola JCM 16345 | 2 591 679 | 65.0 | 1 | 浅海海水 |
| 32 | GCF_001698205.1 | Tsuneonella dongtanensis KCTC 22672 | 3 009 495 | 65.8 | 1 | 浅海沉积物 |
| 33 | GCF_001719165.1 | Erythrobacter litoralis DSM 8509 | 3 251 353 | 65.2 | 1 | 浅海海水 |
| 34 | GCF_001886695.1 | Aurantiacibacter gangjinensis JCM 15420 | 2 724 951 | 62.7 | 2 | 浅海海水 |
| 35 | GCF_002155305.1 | Erythrobacter dokdonensis DSM 17193 | 2 996 446 | 64.8 | 13 | 浅海海水 |
| 36 | GCF_002155425.1 | Erythrobacter donghaensis DSM 16220 | 3 372 281 | 66.2 | 11 | 浅海海水 |
| 37 | GCF_002155655.1 | Erythrobacter sanguineus JCM 20691 | 3 018 761 | 63.6 | 34 | 浅海海水 |
| 38 | GCF_002237615.1 | Qipengyuania flava VG1 | 3 184 307 | 63.3 | 1 | 浅海沉积物 |
| 39 | GCF_002269345.1 | Tsuneonella mangrovi C9-11 | 2 702 851 | 63.5 | 1 | 浅海沉积物 |
| 40 | GCF_002556635.1 | Novosphingobium sp. PC22D | 5 024 522 | 65.8 | 57 | 深海海水 |
| 41 | GCF_002795865.1 | Qipengyuania seohaensis SW-135 | 2 942 673 | 61.7 | 1 | 浅海沉积物 |
| 42 | GCF_002870965.1 | Tsuneonella flava MS1-4 | 3 275 964 | 60.5 | 29 | 浅海沉积物 |
| 43 | GCF_002895025.1 | Erythrobacter sp. SAORIC-644 | 3 549 198 | 60.5 | 25 | 深海海水 |
| 44 | GCF_003264115.1 | Erythrobacter sp. KY5 | 3 311 272 | 60.8 | 1 | 浅海海水 |
| 45 | GCF_003340475.1 | Alteripontixanthobacter maritimus HME9302 | 2 676 898 | 60.8 | 2 | 浅海海水 |
| 46 | GCF_003355455.1 | Erythrobacter aureus YH-07 | 3 372 129 | 62.0 | 2 | 浅海沉积物 |
| 47 | GCF_003363135.1 | Alteriqipengyuania lutimaris S-5 | 3 290 764 | 65.4 | 12 | 浅海海水 |
| 48 | GCF_003569745.1 | Altererythrobacter sp. B11 | 3 842 046 | 65.4 | 1 | 深海沉积物 |
| 49 | GCF_003584015.1 | Aurantiacibacter xanthus CCTCC AB 2015396 | 4 380 877 | 64.5 | 146 | 浅海海水 |
| 50 | GCF_003584125.1 | Aurantiacibacter zhengii V18 | 3 803 809 | 62.7 | 29 | 深海沉积物 |
| 51 | GCF_003605475.1 | Aurantiacibacter aquimixticola JSSK-14 | 2 552 891 | 63.0 | 2 | 浅海海水 |
| 52 | GCF_003605755.1 | Tsuneonella suprasediminis Ery12 | 3 304 416 | 60.9 | 22 | 深海海水 |
| 53 | GCF_004004515.1 | Croceicoccus ponticola GM-16 | 3 244 470 | 62.1 | 17 | 浅海海水 |
| 54 | GCF_004296255.1 | Qipengyuania flava KJ5 | 2 819 202 | 63.9 | 1 | 浅海海水 |
| 55 | GCF_004965515.1 | Alteraurantiacibacter aquimixticola SSKS-13 | 3 433 901 | 63.9 | 5 | 浅海沉积物 |
| 56 | GCF_005434975.1 | Parerythrobacter lutipelagi GH1-16 | 3 097 651 | 60.6 | 2 | 浅海沉积物 |
| 57 | GCF_005884405.1 | Qipengyuania marisflavi KEM-5 | 2 669 685 | 61.7 | 18 | 浅海海水 |
| 58 | GCF_006542605.1 | Porphyrobacter sp. YT40 | 3 943 368 | 65.8 | 2 | 浅海海水 |
| 59 | GCF_007004095.1 | Erythrobacter insulae JBTF-M21 | 2 986 343 | 57.0 | 3 | 浅海海水 |
| 60 | GCF_007570835.1 | Croceicoccus sediminis S2-4-2 | 3 548 496 | 63.0 | 9 | 浅海沉积物 |
| 61 | GCF_007830695.1 | Altererythrobacter ishigakiensis ATCC BAA-2084 | 2 672 716 | 56.9 | 4 | 浅海沉积物 |
| 62 | GCF_008107685.1 | Novosphingobium sp. BW1 | 4 460 758 | 64.5 | 134 | 浅海沉积物 |
| 63 | GCF_008932225.1 | Qipengyuania flava 21-3 | 3 229 668 | 63.6 | 1 | 深海沉积物 |
| 64 | GCF_009746585.1 | Novosphingobium aquimarinum M24A2M | 4 377 191 | 64.0 | 57 | 浅海海水 |
| 65 | GCF_009827295.1 | Qipengyuania pelagi JCM 17468 | 3 031 738 | 64.2 | 9 | 浅海海水 |
| 66 | GCF_009827315.1 | Qipengyuania gaetbuli DSM 16225 | 2 780 525 | 64.1 | 4 | 浅海沉积物 |
| 67 | GCF_009827395.1 | Pontixanthobacter aquaemixtae KCTC 52763 | 2 983 943 | 58.5 | 3 | 浅海海水 |
| 68 | GCF_009827435.1 | Croceibacterium salegens MCCC 1K01500 | 3 636 778 | 64.6 | 63 | 浅海沉积物 |
| 69 | GCF_009827445.1 | Qipengyuania aquimaris JCM 12189 | 2 662 510 | 61.8 | 1 | 浅海海水 |
| 70 | GCF_009827455.1 | Pontixanthobacter aestiaquae KCTC 42006 | 2 868 693 | 57.2 | 2 | 浅海海水 |
| 71 | GCF_009827515.1 | Pelagerythrobacter marinus H32 | 3 001 465 | 68.2 | 16 | 深海海水 |
| 72 | GCF_009827535.1 | Qipengyuania oceanensis MCCC 1A09965 | 2 874 198 | 63.9 | 14 | 深海沉积物 |
| 73 | GCF_009827545.1 | Pontixanthobacter gangjinensis JCM 17802 | 2 888 119 | 55.5 | 1 | 浅海沉积物 |
| 74 | GCF_009827615.1 | Allopontixanthobacter confluentis KCTC 52259 | 2 934 197 | 59.1 | 4 | 浅海海水 |
| 75 | GCF_009827635.1 | Parapontixanthobacter aurantiacus MCCC 1A09962 | 2 895 130 | 61.2 | 7 | 深海沉积物 |
| 76 | GCF_009828015.1 | Qipengyuania citrea CGMCC 1.8703 | 3 031 321 | 64.2 | 24 | 浅海海水 |
| 77 | GCF_009828025.1 | Qipengyuania algicida KEMB 9005-328 | 3 215 496 | 60.7 | 21 | 浅海海水 |
| 78 | GCF_009828095.1 | Pontixanthobacter luteolus SW-109 | 2 887 613 | 59.3 | 3 | 浅海海水 |
| 79 | GCF_009828115.1 | Allopontixanthobacter sediminis KCTC 42453 | 3 158 912 | 61.5 | 6 | 浅海沉积物 |
| 80 | GCF_011762005.1 | Qipengyuania flava DSM 16421 | 2 813 855 | 63.9 | 5 | 浅海海水 |
| 81 | GCF_012972675.1 | Pontixanthobacter rizhaonensis RZ02 | 2 790 898 | 55.5 | 4 | 浅海海水 |
| 82 | GCF_012979275.1 | Pseudopontixanthobacter vadosimaris JL3514 | 2 629 682 | 63.0 | 43 | 浅海海水 |
| 83 | GCF_013260645.1 | Erythrobacter mangrovi EB310 | 3 058 693 | 62.9 | 1 | 浅海沉积物 |
| 84 | GCF_013370205.1 | Actirhodobacter atriluteus HHU K3-1 | 2 939 611 | 62.1 | 23 | 浅海海水 |
| 85 | GCF_013371495.1 | Altererythrobacter lutimaris JGD-16 | 2 952 860 | 57.8 | 33 | 浅海海水 |
| 86 | GCF_013408095.1 | Novosphingobium marinum DSM 29043 | 3 591 024 | 64.2 | 22 | 深海海水 |
| 87 | GCF_014191645.1 | Alteriqipengyuania lutimaris CECT 8624 | 3 269 205 | 65.5 | 6 | 浅海沉积物 |
| 88 | GCF_014199315.1 | Croceibacterium atlanticum DSM 100738 | 3 457 495 | 61.9 | 22 | 深海沉积物 |
| 89 | GCF_014640055.1 | Novosphingobium marinum CGMCC 1.12918 | 3 590 209 | 64.2 | 27 | 深海海水 |
| 90 | GCF_014641655.1 | Alteriqipengyuania_A marina CGMCC 1.15297 | 2 634 902 | 64.9 | 56 | 浅海海水 |
| 91 | GCF_014642495.1 | Croceicoccus pelagius CGMCC 1.15358 | 3 228 892 | 62.8 | 42 | 浅海海水 |
| 92 | GCF_014642595.1 | Croceicoccus mobilis CGMCC 1.15360 | 4 116 752 | 62.6 | 37 | 深海沉积物 |
| 93 | GCF_014645195.1 | Novosphingobium indicum CGMCC 1.6784 | 4 961 652 | 63.0 | 103 | 深海沉积物 |
| 94 | GCF_014705715.1 | Erythrobacter rubeus KMU-140 | 3 042 712 | 60.6 | 4 | 浅海海水 |
| 95 | GCF_015529805.1 | Qipengyuania soli 6D36 | 2 910 144 | 63.3 | 1 | 浅海沉积物 |
| 96 | GCF_015865035.1 | Novosphingobium aureum YJ-S2-02 | 4 342 390 | 65.5 | 39 | 浅海沉积物 |
| 97 | GCF_015999305.1 | Erythrobacter sp. YJ-T3-07 | 3 317 720 | 64.3 | 494 | 浅海沉积物 |
| 98 | GCF_016019885.1 | Aurantiacibacter sediminis JGD-13 | 2 668 619 | 60.2 | 14 | 浅海沉积物 |
| 99 | GCF_016745095.1 | Croceicoccus sp. YJ47 | 3 353 209 | 64.4 | 1 | 深海海水 |
| 100 | GCF_017312445.1 | Novosphingobium sp. PY1 | 4 764 096 | 63.4 | 36 | 浅海沉积物 |
| 101 | GCF_018205975.1 | Erythrobacter sp. JK5 | 3 340 457 | 64.2 | 1 | 浅海沉积物 |
| 102 | GCF_018398395.1 | Croceicoccus gelatinilyticus 1NDH52 | 3 790 455 | 62.6 | 43 | 浅海沉积物 |
| 103 | GCF_018417475.1 | Novosphingobium decolorationis 502str22 | 4 750 375 | 65.5 | 2 | 深海沉积物 |
| 104 | GCF_018636735.1 | Erythrobacter sp. 3-20A1M | 3 116 220 | 64.8 | 1 | 浅海海水 |
| 105 | GCF_900115585.1 | Qipengyuania nanhaisediminis CGMCC 1.7715 | 2 899 438 | 62.0 | 12 | 浅海沉积物 |
| 106 | GCF_900143235.1 | Erythrobacter sanguineus DSM 11032 | 2 978 798 | 63.8 | 25 | 浅海海水 |
| 107 | GCF_900177715.1 | Altererythrobacter xiamenensis CGMCC 1.12494 | 3 082 408 | 61.8 | 5 | 浅海海水 |
基于获取的单拷贝OCs蛋白质序列,使用MAFFT v7软件[17](参数“-auto”)进行序列对齐,并通过trimAL v1.4.1软件[18](参数“-automated1”)对对齐序列进行修剪,以去除冗余序列,然后通过实验室自编的Perl脚本[15]串联氨基酸序列。随后,使用IQ-TREE v1.6.1软件[19](参数“-m MFP”)预测最佳氨基酸替换模型,并继续采用此软件进行最大似然法系统发育分析,抽检值设定为1 000次。获得基因组进化树后,使用Chiplot在线服务(https://www.chiplot.online/tvbot.html)进行可视化、修饰和标注。
1.2 类胡萝卜素合成基因在基因组进化树上的标注
基于Swiss-Prot数据库(https://www.uniprot.org/),获取类胡萝卜素合成基因(crtBEGIWXYZ)序列,并通过BLAST软件构建类胡萝卜素合成基因本地数据库。将基因组注释的赤杆菌科CDSs氨基酸序列与该数据库进行比对,参数为“-evalue 1e-5-num_alignments 1 -max_target_seqs 1”,以筛选序列一致性高于30%的序列,并使用Chiplot在线服务在基因组系统发育树上标注类胡萝卜素合成基因。
1.3 类胡萝卜素合成基因系统发育树的构建
针对不同类胡萝卜素合成基因序列,分别构建最大似然法系统发育树。构建方法与1.1节中基因组系统发育分析方法相近,其中WAG+F+R5为crtB的最佳替换模型,LG+F+R7为crtE的最佳替换模型,pREV+F+R6为crtG的最佳替换模型,WAG+F+R8为crtI的最佳替换模型,LG+F+I+G4为crtW和crtZ的最佳替换模型,LG+F+R6为crtY的最佳替换模型。使用Chiplot在线服务对所有类胡萝卜素合成基因系统发育树进行进一步修饰,标注菌株所属遗传类群并添加图注。后续分析中如有需特别标注的菌株通过Photoshop进行标注。
2 结果与分析
2.1 海洋赤杆菌科菌株基因组分类特征
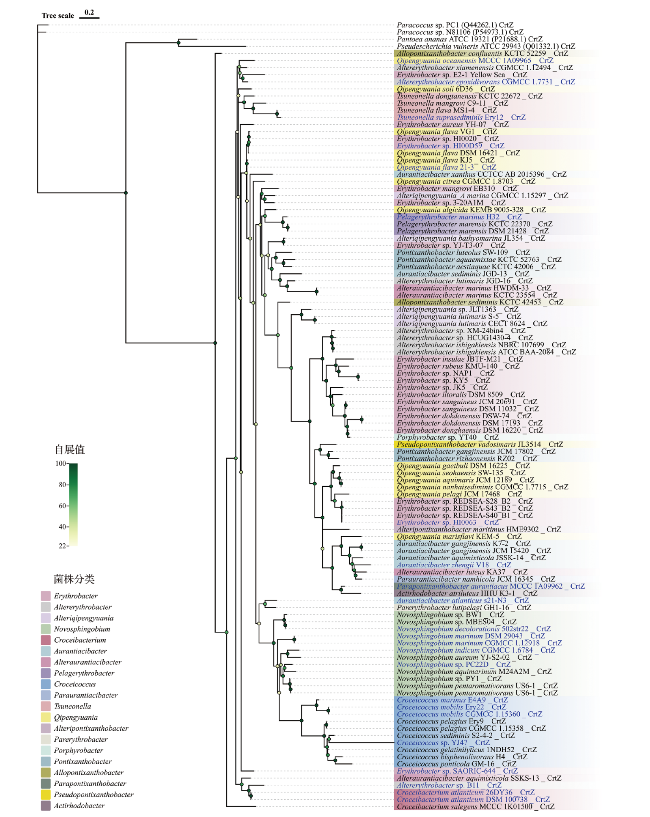
本研究使用107个分离自海洋的赤杆菌科菌株基因组,计算其ANI后发现,ANI>99%的基因组在crt基因的分布中表现出存在和数量上的差异,因此保留这些基因组。通过最大似然法构建系统发育树,标注出20个属,并且可将赤杆菌科划分为九个类群:第一类群由Novosphingobium全部菌株组成,第二类群由Croceicoccus全部菌株组成,第三类群由Croceibacterium、Paraurantiacibacter、Alteraurantiacibacter和Aurantiacibacter的全部菌株以及一个Altererythrobacter菌株组成,第四类群由Pelagerythrobacter和Tsuneonella全部菌株组成,第五类群由Alteripontixanthobacter和Pontixanthobacter全部菌株组成,第六类群由Parerythrobacter全部菌株、Altererythrobacter的7个菌株和Erythrobacter的12个菌株组成,第七类群由Alteriqipengyuania全部菌株和Erythrobacter的2个菌株组成,第八类群由Qipeng-yuania全部菌株和Erythrobacter的9个菌株组成,第九类群由Actirhodobacter和Parapontixanthobacter全部菌株组成(图2)。其中21.5%的菌株分离自深海环境,78.5%的菌株分离自浅海环境(包括海水和沉积物)。
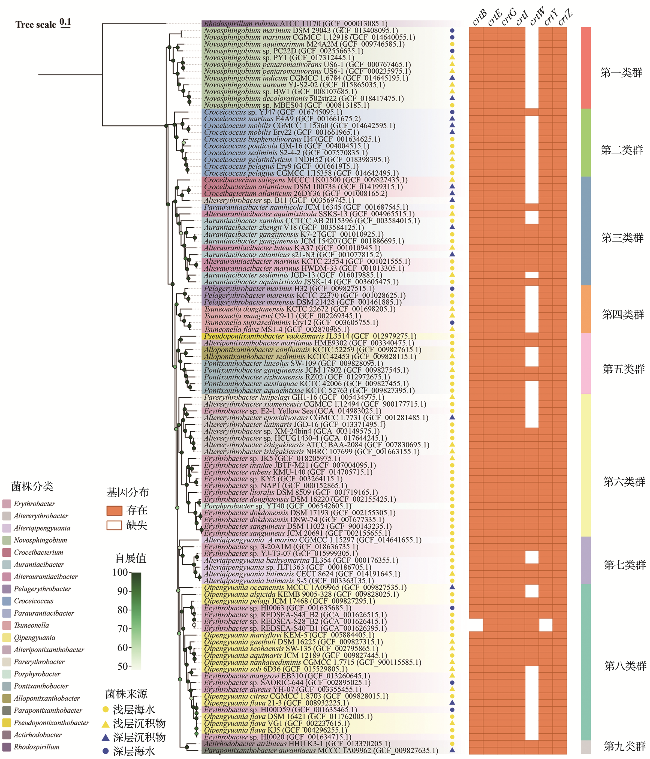
2.2 类胡萝卜素合成基因在赤杆菌科类群中的分布特征
crtEBI基因是所有类胡萝卜素合成途径中必需的基因,可使类胡萝卜素到达进一步合成代谢的分枝点,crtY基因是形成β-胡萝卜素的必需基因,crtGWXZ基因在不同的组合表达下可催化β-胡萝卜素形成多种中间体。不同类胡萝卜素合成基因在赤杆菌科菌株中的存在与分布情况标注并展示在图2中。107个赤杆菌科菌株全部含有crtEIGZ基因,且均不存在crtX基因,表明通过crtGZ基因衍生类胡萝卜素的过程在该科中可能广泛存在。crtBY基因除了在Erythrobacter sp. REDSEA-S40_B1和Erythrobacter sp. REDSEA-S28_B2两个菌株中缺失以外,其余105个菌株都含有crtBY基因,表明这两个菌株不具备完整的类胡萝卜素合成途径。在107个赤杆菌科菌株中,47株含有crtW基因,占总菌株数的43.9%。含有crtW基因的菌株集中于Erythrobacter(69.6%)、Qipengyuania(42.9%)、Altererythrobacter(50%)、Alteriqipengyuania(80%)、Aurantiacibacter(71.4%)、Alteraurantiacibacter(75%)、Paraurantiacibacter(100%)、Alteripontixanthobacter(100%)、Porphyrobacter(100%)、Pontixanthobacter(40%)、Parapontixanthobacter(100%)、Pseudopontixanthobacter(100%)、Actirhodobacter(100%)和Croceicoccus(10%),显示出较大的属间差异。
2.3 类胡萝卜素合成基因在赤杆菌科中的系统发育
crtB基因在海洋赤杆菌科中分布广泛,如图2所示,除了Erythrobacter sp. REDSEA-S40_B1和Erythrobacter sp. REDSEA-S28_B2以外,107株菌中有105株含有crtB基因。且crtB基因在海洋赤杆菌科进化过程中较为保守。由图3所示,在crtB基因的系统发育树中,Qipengyuania oceanensis MCCC 1A09965和Allopontixanthobacter sediminis KCTC 42453表现出与基因组进化树聚类情况的不一致,这两株菌分别分离自深层和浅层沉积物,在来源中没有特异性,推测这两株菌可能通过基因的水平转移获得crtB基因。而其余Novosphingobium、Croceicoccus、Erythrobacter、Aurantiacibacter等同属菌聚类情况与基因组进化树基本一致,说明该科所获得的crtB基因大部分来源于基因的垂直遗传。
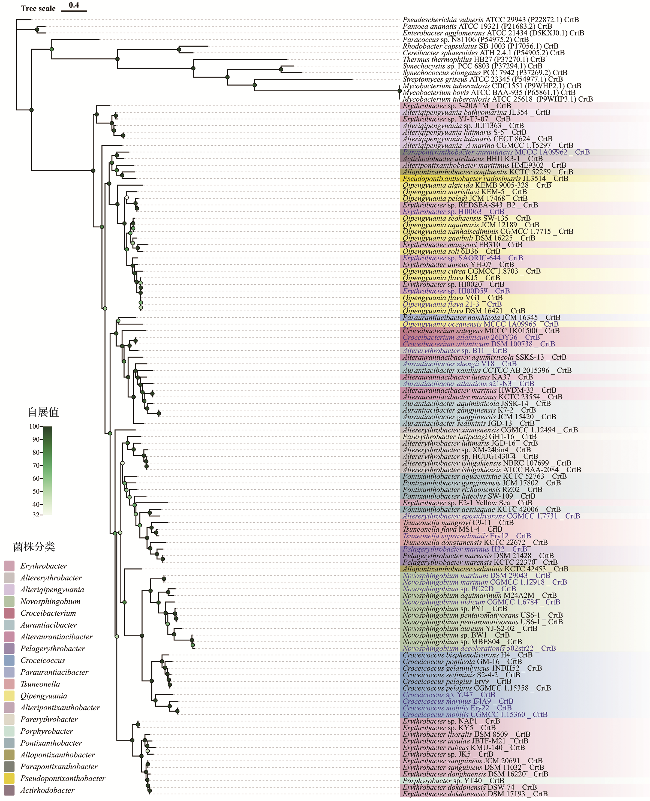
在crtE基因的系统发育树中,菌株Alteriqipengyuania_A marina CGMCC 1.15297与基因组进化树相比表现出聚类的不一致,并且有105个菌株同时含有2个crtE基因,其中Altererythrobacter、Tsuneonella、Pontixanthobacter、Allopontixanthobacter、Erythrobacter、Qipengyuania属内共有7个菌株含有3个crtE基因,表明这些菌株可能通过基因的水平转移获得crtE基因。在具有多个crtE基因的105个菌株中,分离自浅海环境的菌株多于深海环境,但分离自深海环境的23个菌株全部含有多个crtE基因,分离自浅海环境的部分菌株只存在单个crtE基因。
crtI基因在海洋赤杆菌科菌株中表现出很高的丰度。所有107个海洋赤杆菌科菌株都含有2个及以上的crtI基因,其中含有5个及以上crtI基因的菌株占总菌株数的27.1%,集中在Allopontixanthobacter(50%)、Altererythrobacter(50%)、Aurantiacibacter(28.6%)、Croceicoccus(20%)、Erythrobacter(47.8%)、Novosphin-gobium(33.3%)、Parerythrobacter(100%)、Pontixan-thobacter(60%)和Porphyrobacter(100%)。这些含有5个及以上crtI基因的菌株中有89.7%来自浅海环境,有41.4%来自沉积环境,表明浅海环境中含有crtI基因的菌株较多。crtI基因在海洋赤杆菌科菌株中的进化过程更为复杂,且多数菌株聚类情况与基因组进化树不一致,说明该科所获得的crtI基因大部分来源于基因的水平转移。
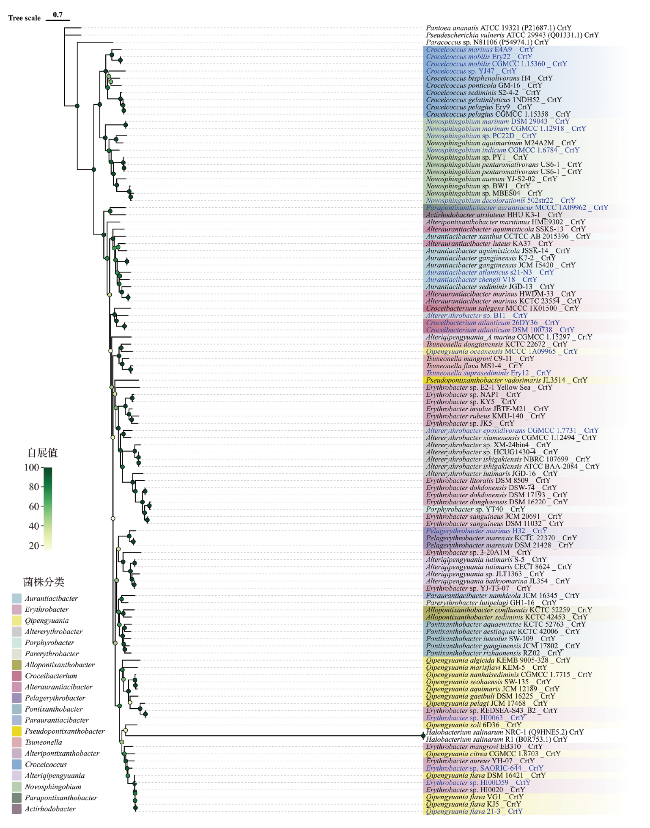
crtY基因可将番茄红素环化形成β-胡萝卜素,其在海洋赤杆菌科中分布广泛,在浅海和深海环境中均有分布。如图2所示,与crtB一致,除了Erythrobacter sp. REDSEA-S40_B1和Erythrobacter sp. REDSEA-S28_B2以外,107株菌中有105株含有crtY基因,表明在海洋赤杆菌科中通常是由crtY基因催化形成β环的类胡萝卜素。crtY基因在进化过程中也较为保守。将其系统发育树中的分支与基因组进化树进行对照,发现除了Alteripontixanthobacter maritimus HME9302、Alteriqipengyuania_A marina CGMCC 1.15297、Qipeng-yuania oceanensis MCCC 1A09965以及Paraurantiaci-bacter namhicola JCM 16345以外,其余菌株的聚类关系与基因组进化树基本一致,而这些不一致的菌株来源广泛,包括深海和浅海环境,这可能意味着crtY基因在海洋赤杆菌科的进化过程中通过垂直转移传递,而与海水深度无关。值得一提的是,Qipengyuania soli 6D36显示与Halobacterium的NRC-1、R1聚类,可能表明该菌株的crtY基因来自Halobacterium的水平转移。
crtZ基因在海洋赤杆菌科中分布也很广泛,107个菌株全部含有crtZ基因(图2),其分布与分离环境并无明显相关性。crtZ基因在各属内的进化过程也较为保守,将系统发育树中的分支与基因组进化树进行对比,发现大部分菌株的聚类与基因组进化树基本一致,表明海洋赤杆菌科中crtZ基因的主要来源为垂直转移(图5)。而Qipengyuania属的MCCC 1A09965、6D36、KEMB 9005-328、KEM-5,Aurantiacibacter属的CCTCC AB 2015396,Altererythrobacter属的JGD-16,这些菌株的聚类关系与基因组进化树不一致,推测其crtZ基因来源于科内的基因水平转移。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
crtG基因在107株菌中也全部存在。通过统计发现,有43株菌含有2个及以上crtG基因,分别隶属于16个属,表明crtG基因在属间分布也十分广泛。与基因组进化树相比较,发现这些含有多个crtG基因的菌株其聚类关系大多发生了改变,表明海洋赤杆菌科细菌在进化过程中通过基因水平转移获得crtG基因。
crtW基因在海洋赤杆菌科中分布较少,107株菌中只有47株菌含有crtW基因,且更趋向分布于浅海环境中,50%的浅海来源菌株含有crtW基因,只有5株来自深海环境,占据所有深海来源菌株的21.7%。其中Croceicoccus属只有来源于深海环境的YJ47菌株含有crtW基因,说明来源于深海的Croceicoccus属菌株更趋向含有crtW基因。根据crtW基因的系统发育树,Erythrobacter sp. NAP1具有2个crtW基因,而Aurantiacibacter atlanticus s21-N3聚类关系与基因组进化树不一致,并且与Haematococcus lacustris聚集在一个分支上,表明这株菌可能通过基因的水平转移获得crtW基因。其余菌株同属菌聚类情况与基因组进化树基本一致,说明该科所获得的crtW基因大部分来源于基因的垂直转移。
3 讨论
3.1 海洋赤杆菌科细菌中类胡萝卜素合成基因的分布情况
根据基因组系统发育树,除了crtW基因以外,类胡萝卜素合成基因在海洋赤杆菌科细菌中广泛存在。海洋赤杆菌科107个菌株中均只含有一个crtBYZ基因,而crtEIGW基因在赤杆菌科菌株中含有多个拷贝,这可能引起系统发育树结构的多系发生,且部分基因不处于类胡萝卜素合成基因簇中,从而不发挥功能。
crtB基因可将两分子的类胡萝卜素前体物质牻牛儿基牻牛儿基焦磷酸(geranylgeranyl pyrophosphate, GGPP,C20)尾尾连接,形成第一个无色的C40类胡萝卜素——八氢番茄红素,是类胡萝卜素合成途径中所必需的酶。随后,八氢番茄红素经crtI基因催化的去饱和反应形成有颜色的番茄红素,其可经环化形成一系列的有1个或2个环的β-环、ε-环或γ-环的类胡萝卜素。而crtY基因可将番茄红素催化形成β-胡萝卜素,也是类胡萝卜素合成途径中重要的酶。β-胡萝卜素可经crtZ和crtW等基因催化形成玉米黄质、角黄素、虾青素等多种中间体[12]。赤杆菌科中Erythrobacter属含有细菌叶绿素和光合作用元件,可以利用光能作为能量补充[2],是典型的AAPB类群,研究发现来自深海环境的该属菌株都不含crtW基因,推测该属菌株独特的光合异养形式与分离环境对其类胡萝卜素合成基因的分布产生了一定影响。此外,除了crtB和crtY基因在Erythrobacter sp. REDSEA-S40_B1和Erythrobacter sp. REDSEA-S28_B2中表现出缺失,导致其类胡萝卜素合成通路不完整以外,其余菌株均含有完整的类胡萝卜素合成途径,而这些菌株来源广泛。因此在本研究中浅海与深海环境下、水环境与沉积环境下类胡萝卜素合成基因的存在与分布并无明显差异,表明在海洋赤杆菌科细菌中,环境不是影响其存在与分布的主要因素,而该科菌株的系统发育与其分布关系密切。
3.2 类胡萝卜素合成基因在海洋赤杆菌科细菌中的进化机制
类胡萝卜素为萜类化合物,在海洋微生物中主要起到光保护,维持细胞膜完整性,缓解低温、高压、高盐等压力及抗氧化的作用[20]。一般来说,微生物对不同的环境条件有其进化适应机制,以发挥正常功能。它们的膜以及遗传产物可以通过特定的刺激发生改变,这种刺激是由于温度、光照、pH、压力和盐度等非生物条件的变化而出现的[21-22]。例如高光照压力和氮源缺乏是诱导Dunaliella salina的β-胡萝卜素积累的重要环境因素[23⇓-25]。但在本研究中,我们发现海洋赤杆菌科细菌所处的深海或浅海的不同环境条件与其类胡萝卜素合成基因产生类型及分布没有明显相关性。虽然深海环境无光照条件,但其他的高压、低温等胁迫也可能使得类胡萝卜素应激产生,因而无法证明海洋赤杆菌科细菌中类胡萝卜素合成基因的存在与分布可通过不同环境条件发生适应改变,而是通过自身基因的水平转移或垂直系统发育等其他潜在的进化适应方式。
细菌可以通过水平基因转移从其他物种获得基因,并将基因分布在不同物种之间[26]。在原核生物中,影响类胡萝卜素合成途径的一个关键因素是水平基因转移和来自共同祖先的异功能同源物的基因家族的进化[27-28]。例如C40类胡萝卜素生物合成所需基因crtP-Qc和crtH,在好氧蓝藻和绿硫厌氧细菌中存在水平基因转移[29]。除了单基因转移外,包括结构性光合作用基因、细菌叶绿素合成基因和类胡萝卜素合成途径的所有crt基因的整个基因簇也可从α-变形菌转移到β-变形菌Rubrivivax中[30]。在本研究中,分别有43个、105个和1个菌株含有2个及以上的crtGEW基因,而107个菌株全部含有两个及以上crtI基因,引起系统发育树结构的多系发生。例如在crtG基因系统发育树中,Novosphingobium sp. PC22D及Novosphingobium aquimarinum M24A2M与Alteriqipengyuania_A marina CGMCC 1.15297聚类,而基于基因组的进化树显示Novosphingobium sp.为一个独立的分支,推测Alteriqipengyuania_A marina CGMCC 1.15297的crtG基因来源为水平转移。Croceicoccus sp.在crtG基因系统发育树中显示出与参考序列Sphingomonas elodea ATCC 31461的聚类关系,也与其在基因组进化树中的独立分支不一致,表明Croceicoccus sp.的crtG基因从亲缘较远的物种水平转移得到。类似的,在crtE基因系统发育树中,Alteriqipengyuania_A marina CGMCC 1.15297与Croceicoccus sp.聚类,在crtY基因系统发育树中,Alteriqipengyuania_A marina CGMCC 1.15297与Tsuneonella sp.聚类,也表明Croceicoccus sp.的crtE基因与Tsuneonella sp.的crtY基因转移到了Alteriqipengyuania_A marina CGMCC 1.15297中。由此推测Alteriqipengyuania_A marina CGMCC 1.15297菌株的crtGEY基因可能是通过其他细菌基因组的引入或重新组合,甚至可能是通过环境DNA的摄取从而进行了水平转移。结合对上述菌株的分析,由于赤杆菌科细菌中具有较多长度不同的同源序列,其较近的亲缘关系使得水平基因转移更容易发生,外源DNA更容易插入细菌基因组或导致细菌中原有的DNA被删除[31],即表现为基因的增加或缺失,从而导致在赤杆菌科细菌中crt基因的分布与数量产生不同。
将基因组系统发育树与不同的类胡萝卜素合成基因的系统发育树进行对照,发现Novosphingobium、Croceicoccus、Tsuneonella、Aurantiacibacter、Pontixanthobacter等属在拓扑结构中表现出一致性,表明其获得的类胡萝卜素合成基因大概率来源于基因的垂直传递。其中,Novosphingobium、Croceicoccus属内的全部物种在基于基因组数据的系统发育树和基于类胡萝卜素合成基因的系统发育树中都分别位于一个独立的分支上,因而这两个属是分类地位明确的属,同时这两个属内的菌株在深、浅海环境中均有分离获得,因此可以较为清晰地表明这两个属内菌株的类胡萝卜素合成基因在其进化过程中发生了垂直传递,而并非环境引起的生态型适应。Erythrobacter sp. REDSEA-S40_B1和Erythrobacter sp. REDSEA-S28_B2均分离自浅海环境,而它们在基于基因组系统发育树和类胡萝卜素合成基因的系统发育树中的聚类关系在除crtBY以外的其他类胡萝卜素合成基因中都保持一致,推测两种菌株中crtBY基因的缺失可能是由于基因在垂直传递的演化过程中丢失,而与环境无关。
根据研究结果,类胡萝卜素合成基因在赤杆菌科菌株中广泛存在,除了约98.1%的菌株含有crtBY基因,约43.9%的菌株中存在crtW基因以外,所有菌株都含有crtEIGZ基因。从不同的海洋来源进行分析,这些菌株未表现出深海或浅海环境对于类胡萝卜素合成基因存在类型的偏好性,由此推测深海与浅海环境并不具备特异性。部分菌株在基于基因组系统发育树和类胡萝卜素合成基因的系统发育树中始终处于独立分支且拓扑结构未发生改变,表明这部分菌株的类胡萝卜素合成基因与系统发育相关;部分菌株表现出系统发育树拓扑结构的不一致,推测在属间存在水平基因转移。
3.3 产类胡萝卜素赤杆菌科微生物资源的应用
类胡萝卜素具有很强的抗氧化、修复、抗炎等作用,其在营养保健品和化妆品等市场有很高的应用价值[32]。目前,人们对健康的日益关注增长了天然类胡萝卜素产品的市场需求,海洋微生物已成为当下生产天然类胡萝卜素的潜在资源[33]。而赤杆菌科微生物含有完整的类胡萝卜素合成通路,并且具有明显的色素积累。从海洋环境中分离出的Altererythrobacter、Erythrobacter、Qipengyuania、Tsuneonella等多个属的菌株可合成具有强抗氧化作用的玉米黄质、虾青素、角黄素、金盏花黄质、海胆酮等多种天然类胡萝卜素[7,34⇓-36],这些新的和稀有的海洋类胡萝卜素在营养保健品、药妆产品的研发中有一定潜力[37]。因此,海洋赤杆菌在医药保健、化妆品等领域中具有很好的应用前景。
4 结论与展望
本研究通过基于类胡萝卜素合成基因和基因组数据的系统发育分析,揭示了海洋赤杆菌科细菌中类胡萝卜素合成基因的存在与分布情况,并探讨了类胡萝卜素合成基因在海洋赤杆菌科菌株中的进化机制。
研究表明海洋来源的赤杆菌科菌株均含有crtEIGZ基因,约98.1%的菌株含有crtBY基因,约43.9%的菌株含有crtW基因,但其类胡萝卜素合成基因与环境因素无直接关联性。Novosphingobium、Croceicoccus、Erythrobacter、Aurantiacibacter等同属菌株在crtBYZW基因中表现出其类胡萝卜素合成基因与系统发育相关,而在crtEIG基因中多表现出与系统发育树拓扑结构的不一致,推测在属间存在水平基因转移,这也为解释其他分布广泛的细菌中类胡萝卜素的合成途径及基因分布提供了参考。然而原核生物中类胡萝卜素在各种环境条件下存在着相应的应激作用[38],暂时还未有研究分析原核生物中类胡萝卜素合成基因与不同的环境胁迫之间的关联,目前的研究主要聚焦于类胡萝卜素合成基因的进化来源,未来可针对类胡萝卜素在原核生物中产生的不同应激机制来探究其合成基因的分布。