
0 引言
深海是地球上最大的生态系统,其生境复杂独特,孕育了丰富多样的海洋生物[1]。深海生态系统规模庞大,人类对深海生物群落的食物来源、能量传递等过程的了解不够深入。因此,开展相关研究是认识深海生物多样性维持机制的关键之一,可为可持续管理提供重要支撑[2]。大量研究显示,上层海洋浮游植物产生的有机物是深海生物群落的主要食物来源[3-4]。但绝大部分有机质在沉降过程中,在1 000 m以上的水层即被矿化或吸收,仅余1%~3%难以降解的颗粒有机质可到达深海[5]。深海的食物资源极度匮乏,不同生物类群展现出不同的食物偏好。例如,一些深海鱼类以垂直迁移能力较强的大型浮游动物和中深层水中的虾为主要捕食对象[6],偶尔以其他底栖生物作为其食物来源[7];而滤食性生物一般利用水体中不同类型的颗粒物作为其食物来源[8]。尽管目前对深海生物食物来源的研究已取得了一定进展,但对深海底栖生物的食物来源和营养关系的认识仍然非常有限。
海山通常是指深海大洋中位于水面以下、高度大于1 000 m的隆起地形,是一种典型的深海生境。据报道,全球深海海山超过15万座[13],崎岖的地形和特殊的水文特征共同孕育出独特的深海海山生态系统[14]。LAVELLE等[15]的研究表明,海山的存在增强了水体垂直混合和局部海洋流动,同时在海山尾迹中产生涡流。这些变化有助于提高海山周围的初级生产力,为海山生境拥有更多的物种和更高的生物量提供了可能,使海山成为生产力、生物量和生物多样性研究的热点区域[16]。尽管海山生态系统的重要性日益凸显,但关于深海海山底栖生物群落的食物来源和营养结构等方面的认识仍十分有限。有研究发现悬浮颗粒物和缓慢下沉的有机颗粒是巨型底栖生物的直接食物来源[17],但不同生物类群在食物选择方面可能存在显著差异。
维嘉平顶海山(Weijia Guyot)位于西北太平洋麦哲伦海山群南端,是中国富钴结壳勘探合同区内的海山。这一海山位于远离大陆的热带/亚热带寡营养西北太平洋,上层水体中的初级生产力较低。目前对西北太平洋深海海山巨型底栖生物食物来源等问题的研究较少。SHEN等[18]研究发现,维嘉海山底栖生物群落以海绵、珊瑚、海百合和蛇尾等巨型底栖生物为主。另有研究表明,深海生境中,海绵多为滤食性生物,常具独特的身体形状,以便更容易地获取食物[19-20]。珊瑚是食性较广泛的消费者,能够以颗粒有机质、悬浮沉积物、细菌、浮游动物和浮游植物为食[21-22]。海百合和蛇尾属于棘皮动物门,是古老而特殊的海洋生物[23]。海百合是滤食性生物,通过变动腕和羽枝的位置对水流中的碎屑和浮游生物进行滤食[24-25]。蛇尾可以生活在底泥表面或者藏匿在泥下,也可以附在珊瑚、海绵等固着物上[26];它们的食物来源广泛,包括浮游植物、浮游动物、多毛类动物、碎屑及腐肉,其食性主要为滤食性和腐食性[27]。本研究分析了维嘉海山巨型底栖生物、浮游动物和表层沉积物的碳、氮稳定同位素组成,旨在揭示维嘉海山不同底栖生物类群的食物来源,促进对深海海山底栖生物群落营养结构的认识。
1 材料与方法
1.1 样品采集
本研究利用大洋61、76和80航次在西北太平洋维嘉海山采集的底栖生物、浮游动物和表层沉积物样品开展研究,具体采样站位如图1所示。底栖生物样品由“海龙Ⅲ”ROV和“蛟龙号”载人潜水器采集,样品采集后于-20 ℃冷冻保存并带回实验室用于同位素分析。浮游动物样品使用浮游生物分层拖网(MultiNet,HYDRO-BIOS)采集(网衣孔径200 μm),采样深度分别为:0~50 m、50~100 m、100~150 m、150~200 m、200~300 m、300~500 m、500~750 m和750~1 000 m。采集的浮游动物样品用450 °C预处理过的GF/F滤膜(Whatman)过滤,-20 ℃冷冻保存,带回实验室用于同位素分析。沉积物样品由“蛟龙号”载人潜水器所携带的插管(push-core)采集,取表层沉积物样于-20 ℃冷冻保存至实验室处理(仅进行15N/14N分析)。
1.2 样品预处理
取适量底栖生物样品,经55 ℃恒温烘干48 h后,用玛瑙研钵研磨成均匀粉末,冷却至室温后分成两份。一份用于分析样品的δ15N值;另一份用稀盐酸(1 mol/L)处理以去除碳酸盐,然后转移至用450 ℃预处理过的GF/F滤膜上,并用去离子水清洗,除去残留的盐酸,之后在55 ℃条件下恒温烘干48 h,用于分析样品的δ13C值。表层沉积物样品用玛瑙研钵研磨成均匀粉末,经50 ℃恒温烘干48 h后,用于分析样品的δ15N值。
对于浮游动物样品,将含有浮游动物的滤膜在55 ℃条件下恒温烘干48 h,冷却至室温后分成两份。一份用于分析样品的δ15N值;另一份用浓盐酸(12 mol/L)酸熏48 h后,用去离子水清洗样品,去除其中残留的盐酸,之后在55 ℃条件下恒温烘干48 h,用于分析样品的δ13C值。
1.3 稳定同位素分析
取适量经过预处理的样品,在锡舟中压实,使用253 Plus型稳定同位素比质谱仪和Conflo IV型连续流接口装置(Thermo Scientific,美国)进行稳定同位素组成分析。碳、氮稳定同位素组成以δ表示(‰):
$ \delta_{\text {sample }}=\left(\frac{R_{\text {sample }}}{R_{\text {standard }}}-1\right) \times 1000$
式中:δsample代表样品的δ15N或δ13C值,Rsample代表实测样品的15N/14N或13C/12C比值,Rstandard表示标准物质的15N/14N或13C/12C比值。稳定碳、氮同位素比值参考的标准物质分别为空气中的氮气(N2)和箭石标准物质(Vienna Pee Dee Belemnite,VPDB)。为确保测试仪器的稳定和数据的质量,测试样品时插入标准品进行分析,每间隔10份样品插入1份标准品进行漂移分析,标准品为USGS40(δ15NAir =-4.52‰,δ13CVPDB=-26.39‰)。δ15N和δ13C的稳定同位素测量相对标准偏差均小于0.3‰。
2 结果
表1 维嘉海山巨型底栖生物各类群的稳定同位素(δ13C和δ15N)值Tab.1 Stable isotope (δ13C and δ15N) values of megabenthos in the Weijia Guyot |
| 底栖生物 | 站位 | 水深/m | δ13C/‰ | δ15N/‰ |
|---|---|---|---|---|
| 海绵Porifera | ||||
| 勾棘海绵Uncinateridae | S04 | 1 907 | -20.5 (0.1) | 15.0 |
| 绢网海绵Farreidae | S04 | 1 796 | -20.6 (0.2) | 18.0 (0.1) |
| 绢网海绵Farrea sp. | S04 | 1 796 | -20.3 (0.1) | 17.8 (0.1) |
| 长茎海绵Caulophacus sp. | ROV11 | 2 380 | -19.1 | 18.1 (0.1) |
| 长茎海绵Caulophacus sp. | S04 | 1 947 | -20.9 (0.1) | 17.3(0.2) |
| 长茎海绵Caulophacus sp. | S06 | 1 681 | -20.7 | 17.3 |
| 沃尔特海绵Walteria sp. n. HC-2 021 | S04 | 1 850 | -21.7 (0.1) | 12.5 (0.1) |
| 沃尔特海绵Walteria sp. n. HC-2 022 | S06 | 1 702 | -21.9 (0.1) | 11.7 (0.1) |
| 沃尔特海绵Walteria sp. | S10 | 1 941 | -21.6 (0.1) | 13.8 (0.1) |
| 黄绿茎球海绵Flavovirens sp. | S05 | 2 193 | -20.3 | 19.2(0.3) |
| 平均值 | -20.8±0.9 | 16.1±2.9 | ||
| 珊瑚Anthozoa | ||||
| 海笔Protoptilum sp. | S08 | 1 776 | -21.1 | 11.6 (0.2) |
| 半红珊瑚hemicorallium sp. | S08 | 2 076 | -15.1 (0.1) | 14.7 (0.1) |
| 拟柳珊瑚Paragorgia sp. | S04 | 1 796 | -20.0 (0.2) | 12.5 (0.1) |
| 金柳珊瑚Chrysogorgia sp. | S04 | 1 796 | -19.6 | 10.4 (0.1) |
| 金柳珊瑚Chrysogorgia sp. | S04 | 1 796 | -21.0 (0.2) | 9.6 (0.1) |
| 金柳珊瑚Chrysogorgia sp. | S05 | 1 791 | -19.7 (0.2) | 10.1 |
| 角柳珊瑚Keratoisididae | S04 | 1 796 | -21.6 (0.1) | 12.0 (0.1) |
| 角柳珊瑚Keratoisididae | S04 | 1 806 | -21.2 (0.1) | 11.9 (0.2) |
| 角柳珊瑚Keratoisididae | S10 | 1 753 | -22.9 | 13.1 (0.2) |
| 裂黑珊瑚Schizopathidae | S06 | 1 885 | -20.6 (0.1) | 14.5 (0.1) |
| 平均值 | -20.3±2.1 | 12.0±1.7 | ||
| 钩虾Gammaridea | ||||
| 隐首钩虾Stegocephalidae | S137 | 1 985 | -19.3 (0.2) | 9.5 (0.2) |
| 隐首钩虾Stegocephalidae | S137 | 1 985 | -19.5 (0.2) | 9.5 (0.2) |
| 隐首钩虾Stegocephalidae | S137 | 1 985 | -19.8 (0.2) | 8.3 (0.3) |
| 平均值 | -19.5±0.3 | 9.1±0.7 | ||
| 海星Asteroidea | ||||
| 奇板海星Colpaster patricki | S08 | 2 070 | -21.0 (0.1) | 14.3 (0.1) |
| 奇板海星Colpaster patricki | S04 | 1 728 | -20.4 | 15.6 |
| 平均值 | -20.7±0.4 | 15.0±0.9 | ||
| 海百合Crinoidea | ||||
| 奇异羽枝海百合Thaumatocrinus sp.2 | ROV11 | 2 380 | -18.7 (0.2) | 15.8 (0.2) |
| 奈氏奇异羽枝海百合Thaumatocrinus naresi | ROV12 | 1 727 | -19.2 (0.3) | 12.6 (0.3) |
| 奈氏奇异羽枝海百合Thaumatocrinus naresi | ROV12 | 1 702 | -22.2 (0.1) | 11.6 (0.1) |
| 海羊齿海百合Antedonidae gen sp.5 | ROV12 | 1 702 | -18.8 (0.1) | 13.5 (0.3) |
| 海羊齿海百合Antedonidae gen sp.5 | S06 | 1 728 | -21.0 (0.1) | 13.8 (0.1) |
| 海羊齿海百合Antedonidae gen sp.8 | S06 | 1 728 | -19.7 (0.2) | 13.9 (0.1) |
| 海羽枝海百合Thalassometridae gen. sp.2 | S04 | 1 796 | -21.3 (0.1) | 13.3 (0.2) |
| 海羽枝海百合Thalassometridae gen. sp.2 | S04 | 1 796 | -20.8 | 12.0 (0.1) |
| 海羽枝海百合Thalassometra sp. | S06 | 1 728 | -20.6 | 13.8 (0.1) |
| 吉列海百合Guillecrinus cf. neocaledonicus | S08 | 2 101 | -20.9 (0.1) | 13.6 (0.1) |
| 吉列海百合Guillecrinus cf. neocaledonicus | S10 | 1 812 | -21.4 (0.1) | 12.9 (0.2) |
| 穹羽枝海百合Sarametra sp. | S04 | 1 947 | -20.8 (0.1) | 11.1 (0.1) |
| 穹羽枝海百合Sarametra sp. | S06 | 1 681 | -22.5 (0.2) | 10.4 (0.1) |
| 平均值 | -20.6±1.2 | 12.9±1.4 | ||
| 蛇尾Ophiuroidea | ||||
| 德式柱蛇尾Ophiocamax drygalskii | ROV12 | 1 752 | -18.3 (0.1) | 15.5 (0.1) |
| 护盾砖蛇尾Ophioplinthaca defensor | ROV12 | 1 702 | -18.8 (0.1) | 12.2 (0.2) |
| 护盾砖蛇尾Ophioplinthaca defensor | ROV12 | 1 702 | -17.3 (0.3) | 13.7 (0.3) |
| 护盾砖蛇尾Ophioplinthaca defensor | ROV12 | 1 702 | -17.9 (0.2) | 13.0 (0.3) |
| 护盾砖蛇尾Ophioplinthaca defensor | S11 | 1 608 | -18.2 (0.2) | 12.0 |
| 护盾砖蛇尾Ophioplinthaca defensor | S11 | 1 608 | -18.2 (0.1) | 13.6 |
| 护盾砖蛇尾Ophioplinthaca defensor | S08 | 2 070 | -18.8 (0.1) | 13.6 (0.1) |
| 护盾砖蛇尾Ophioplinthaca defensor | S08 | 2 070 | -18.4 (0.2) | 14.0 (0.3) |
| 星蛇尾Asteroschema sp. | S08 | 2 076 | -18.5 (0.2) | 16.3 (0.1) |
| 星蛇尾Asteroschema sp. | S04 | 1 796 | -17.7 (0.1) | 15.2 (0.2) |
| 美丽莱拉蛇尾Ophioleila elegans | S04 | 1 850 | -20.6 (0.1) | 12.2 (0.2) |
| 美丽莱拉蛇尾Ophioleila elegans | S04 | 1 850 | -20.8 (0.2) | 12.2 |
| 美丽莱拉蛇尾Ophioleila elegans | S04 | 1 850 | -20.7 (0.1) | 12.9 (0.1) |
| 平均值 | -18.8±1.2 | 13.6±1.4 | ||
| 海参Holothuroidea | ||||
| 帕劳底游参Benthodytes palauta | S05 | 2 525 | -18.1 (0.2) | 13.1 (0.2) |
| 深海参Benthogone sp. | S10 | 1 820 | -19.1 | 13.3 |
| 平均值 | -18.6±0.7 | 13.2±0.1 | ||
| 被囊类Tunicate | ||||
| 被囊类Tunicate | S08 | 2 101 | -18.9 | 17.9 (0.1) |
| 被囊类Tunicate | S08 | 2 157 | -19.2 (0.1) | 18.4 (0.2) |
| 平均值 | -19.1±0.2 | 18.2±0.4 |
注:括号中数字为重复测量的标准差,加粗字体为平均值。 |
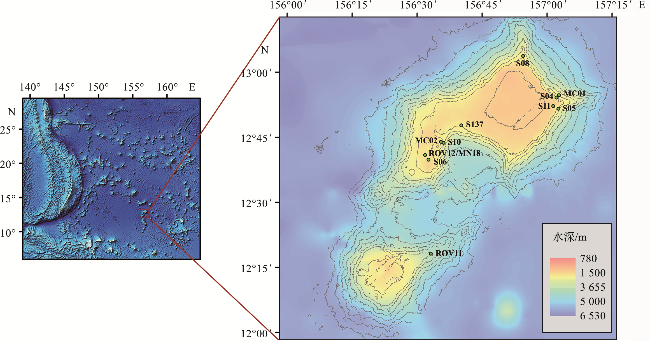
海绵〔包含勾棘海绵科Uncinateridae、绢网海绵科Farreidae、花骨海绵科Rossellidae(长茎海绵Caulophacus sp.)和偕老同穴科Euplectellidae(沃尔特海绵Walteria sp.、黄绿茎球海绵Flavovirens sp.)〕δ13C值的变化范围是-21.9‰~-19.1‰,平均值为(-20.8±0.9)‰;δ15N值的变化范围是11.7‰~19.2‰,平均值为(16.1±2.9)‰。
珊瑚〔包含原羽海鳃科Protoptilidae(海笔Protoptilum sp.)、红珊瑚科Coralliidae(半红珊瑚hemicorallium sp.)、金柳珊瑚科Chrysogorgiidae、角柳珊瑚科Keratoisididae和裂黑珊瑚科Schizopathidae〕δ13C值的变化范围是-22.9‰~-15.1‰,平均值为(-20.3±2.1)‰;δ15N值的变化范围是9.6‰~14.7‰,平均值为(12.0±1.7)‰。
钩虾δ13C值的变化范围是-19.8‰~-19.3‰,平均值为(-19.5±0.3)‰;δ15N值的变化范围是8.3‰~9.5‰,平均值为(9.1±0.7)‰。
棘皮动物包括海星、海百合、蛇尾和海参四类。海星δ13C值的变化范围是-21.0‰~-20.4‰,平均值为(-20.7±0.4)‰;δ15N值的变化范围是14.3‰~15.6‰,平均值为(15.0±0.9)‰。海百合δ13C值的变化范围是-22.5‰~-18.7‰,平均值为(-20.6±1.2)‰;δ15N值的变化范围是10.4‰~15.8‰,平均值为(12.9±1.4)‰。蛇尾δ13C值的变化范围是-20.8‰~-17.3‰,平均值为(-18.8±1.2)‰;δ15N值的变化范围是12.0‰~16.3‰,平均值为(13.6±1.4)‰。海参δ13C值的变化范围是-19.1‰~-18.1‰,平均值为(-18.6±0.7)‰;δ15N值的变化范围是13.1‰~13.3‰,平均值为(13.2±0.1)‰。
被囊类δ13C值的变化范围是-19.2‰~-18.9‰,平均值为(-19.1±0.2)‰;δ15N值的变化范围是17.9‰~18.4‰,平均值为(18.2±0.4)‰。
表2 浮游动物的稳定同位素(δ13C和δ15N)值Tab.2 Stable isotope (δ13C and δ15N) values of zooplankton |
| 站位 | 水深/m | δ13C/‰ | δ15N/‰ |
|---|---|---|---|
| MN18 | 0~50 | -22.1 | 4.9 |
| MN18 | 50~100 | -21.5 | 5.8 |
| MN18 | 100~150 | -21.3 | 6.6 |
| MN18 | 150~200 | -22.0 | 5.6 |
| MN18 | 200~300 | -21.6 | 8.1 |
| MN18 | 300~500 | -23.1 | 8.8 |
| MN18 | 500~750 | -22.4 | 10.2 |
| MN18 | 750~1 000 | -23.0 | 10.5 |
| 平均值 | -22.1±0.7 | 7.6±2.2 |
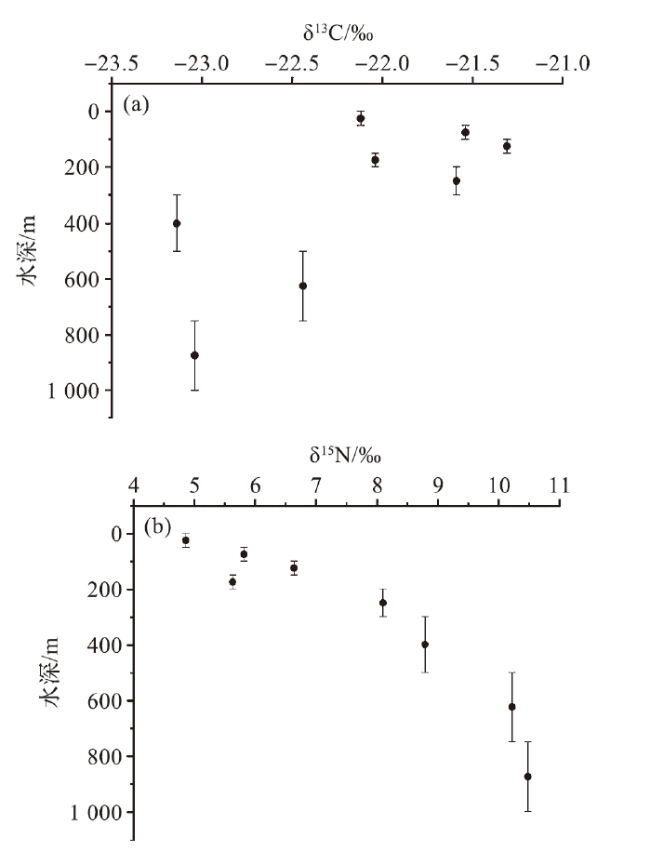
图3 浮游动物δ13C (a) 和 δ15N (b)的垂直分布Fig.3 Vertical distribution of δ13C (a) and δ15N (b) for zooplankton |
表3 表层沉积物的稳定同位素(δ15N)值Tab.3 Stable isotope (δ15N) values of surface sediments |
| 站位 | 水深/m | δ15N/‰ |
|---|---|---|
| MC01 | 2 351 | 7.7(0.2) |
| MC02 | 2 512 | 7.7(0.1) |
| 平均值 | 7.7 |
3 讨论
3.1 浮游动物δ15N值的垂直分布
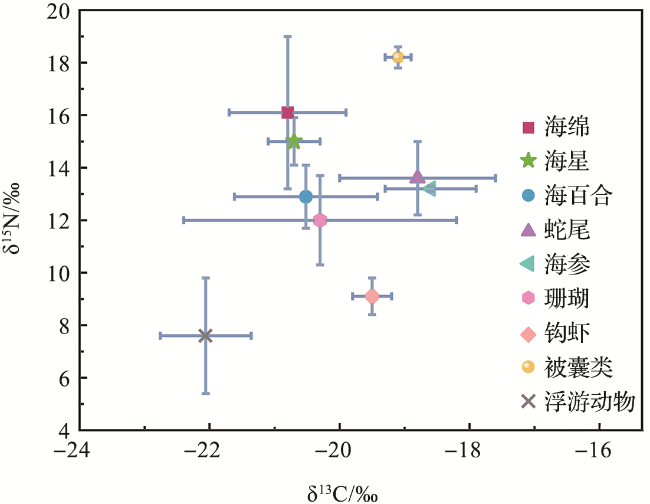
浮游动物的δ15N值表现出随深度增加而增加的趋势(表2和图3),从表层到中深层增加了约6‰。这一现象在前人的研究中也普遍存在,如在西北太平洋采薇海山,浮游动物δ15N值从表层到中深层(0~1 000 m)增加了约5‰[17];在东大西洋、北太平洋和地中海也观测到浮游动物δ15N值具有相似的垂直分布特征[28-31]。这样的变化可能与颗粒有机质的消耗和浮游动物群落结构垂直变化有关。有研究表明,颗粒有机质在沉降过程中,较轻的氮同位素被优先降解,因此输送到深海区的颗粒有机质会富集更多难降解的、较重的氮同位素[29]。深海浮游动物摄食了这些具有较高δ15N值的颗粒物,是导致其δ15N值随水深增大而升高的原因之一。浮游动物群落结构具有显著的垂直变化,不同食性的浮游动物类群在不同水深段的占比不同,食性的差异也表现在稳定同位素值上。有研究发现与植食性和滤食性浮游动物相比,肉食性和杂食性浮游动物的氮同位素值更高[5,8,32-33]。YAMAMAE等[34]对西北太平洋及其边缘海域0~3 000 m浮游动物的垂直分布进行研究,发现杯口水蚤目Poecilostomatoida能够附着在海雪上,以碎屑为食;也有研究发现杯口水蚤目能通过附着在尾海鞘脱落的身体结构上摄取营养,且它们是深水区(250~1 000 m)占比较高的物种[35]。在白令海峡,滤食性浮游动物(主要为剑水蚤科)在表层占比较高,而在深水区占比较低;肉食性浮游动物的分布深度比碎屑食性浮游动物更深[36]。MATTHEWS等[37]对东北太平洋不同浮游动物摄食类型(植食性、肉食性和杂食性)的研究发现,肉食性浮游动物和杂食性浮游动物的丰度峰值出现在更深的水深(100~300 m),而植食性浮游动物的丰度峰值都保持在0~150 m范围内。在缺乏浮游植物、食物有限的深海中,杂食性和肉食性浮游动物类群出现的概率更高,深度是影响海洋中浮游动物分布的最重要因素[38],深海浮游动物群落具有明显的水深分布梯度[39]。因此,浮游动物群落结构变化也可能是其δ15N值垂直变化的重要原因。
3.2 深海海山底栖生物的食物来源
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
浮游动物是底栖食物网中物质与能量传输的重要一环[8]。有研究显示浮游动物是珊瑚等底栖生物的食物来源[43]。维嘉海山750~1 000 m 水深,浮游动物的δ15N值为10.5‰,较该区域巨型底栖生物类群δ15N平均值(13.6‰)低3.1‰,其差值接近1个营养级的富集预期值(3.4‰[44])。这表明浮游动物可能是维嘉海山部分底栖生物类群的食物来源之一。就不同类群而言,沃尔特海绵(S10站)、海羊齿海百合(S06站)、海羽枝海百合(S06站)和部分护盾砖蛇尾的氮同位素值与深层浮游动物氮同位素值差异接近1个营养级的富集值(表1和表2)。因此,深层浮游动物可能是这些底栖生物的重要食物来源。这些生物均是滤食性生物,海绵通过过滤海水中的颗粒物获取食物,海百合和蛇尾通常附着于海绵和珊瑚上,利用其羽枝和腕捕获海水中的颗粒物来摄食[19-20,24-27,45]。目前还缺乏相关研究提供直接证据证明这些底栖生物可以直接利用深水浮游动物,未来应针对这些生物类群的胃含物进行分析,进一步验证它们之间的摄食关系。
沉积物可以在数月至数年的时间尺度上储存有机质,从而为底栖生物提供一个长期的食物来源[46-47]。维嘉海山表层沉积物的δ15N平均值为7.7‰,这与部分巨型底栖生物(沃尔特海绵Walteria sp. n. HC-2022、奈氏奇异羽枝海百合Thaumatocrinus naresi、穹羽枝海百合Sarametra sp.和海笔Protoptilum sp.)的δ15N值相差接近1个营养级。如前文所述,海绵、珊瑚和海百合通常是滤食性生物,通过过滤海水中的悬浮颗粒物获取食物。海山具有特殊的水动力过程,能够引起表层沉积物再悬浮[15,48],这使得滤食性底栖生物摄食海底表层沉积物中的有机物成为可能。JONSSON等[49]的研究也发现巨型底栖生物长期以沉积物作为直接的食物来源。例如海参是典型的沉积食性生物,它们主要通过体表的触手摄取海底表层沉积物中的有机物作为其食物来源[50]。但本研究中海参类的δ15N值(13.2‰)与表层沉积物δ15N值的差异超过1个营养级。这可能与海参摄食了次表层沉积物中的有机质有关。BERGMANN等[32]对弗拉姆海峡的研究也表明,次表层沉积物中含有较高的δ15N值,该海域深海海参较高的δ15N值可能与其进食了次表层沉积物而非新沉降的表层颗粒物有关。此外,有研究表明深海海参可能直接将细菌作为食物来源,或者可能间接利用细菌摄取它们无法从其他途径获得的营养[51]。
4 结论
本研究对西北太平洋维嘉海山的8个底栖生物类群(海绵、珊瑚、钩虾、海星、海百合、蛇尾、海参和被囊类)以及2种潜在的食物来源(浮游动物和表层沉积物)的碳、氮稳定同位素进行了分析研究。结果表明,浮游动物稳定氮同位素值具有明显的随水深增加而升高的趋势;底栖生物δ13C值和δ15N值的变化范围较大,且δ13C和δ15N之间没有显著的相关性;浮游动物和沉积物与部分底栖生物的δ15N差值达到1个营养级的氮同位素富集值,说明它们可能是海山底栖生物群落的重要食物来源。此外,较大的δ13C变化范围意味着底栖生物可能利用了多种不同来源的食物。
本研究分析了深海海山主要底栖生物群落的碳、氮稳定同位素值和潜在食物来源,研究结果为认识深海海山生态系统食物链提供了重要的基础信息,有助于进一步完善对深海海山底栖生物群落营养结构的认识。未来仍需要加强不同食性生物类群和其他食物来源(如悬浮颗粒物、深层沉积物等)的样品采集与研究,以加深对深海海山底栖生态系统物质循环和能量流通途径的理解。