
0 引言
1 深水沉积过程分类及其碳埋藏效应
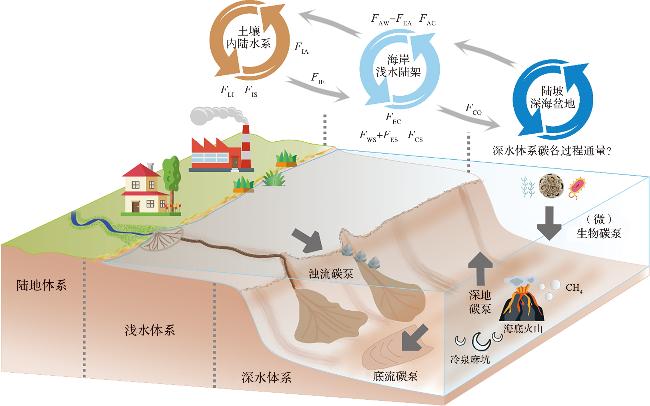
图1 海洋沉积有机质连续输运体系模式图(深海沉积有机质迁移埋藏由(微)生物碳泵、浊流碳泵、底流碳泵、深地碳泵四者共同主导,并与浅水体系、陆地体系耦合,形成“1+2”格局的海洋沉积有机质连续输运体系。陆地体系:土壤-内陆水系有机质连续输运体系;浅水体系:陆地-海岸-浅水陆架有机质连续输运体系;深水体系:陆架边缘-陆坡-深海盆地有机质连续输运体系。FLI:土壤至内陆水系有机碳迁移通量;FIS:内陆水系有机碳埋藏通量;FIA:内陆水系有机碳逸散通量;FIE:内陆水系至海岸有机碳迁移通量;FEC:海岸至浅水陆架有机碳迁移通量;FWS+FES:海岸有机质埋藏通量; FAW-FEA:海岸有机碳吸收通量;FCS:陆架有机质埋藏通量;FAC:陆架水体有机碳吸收通量;FCO:陆架至深海有机碳迁移通量[10]。) Fig.1 Schematic diagram of the continuous transport system of marine sedimentary organic matter (The transport and burial of deep-sea sedimentary organic matter are governed by four synergistic pumps: the biological & microbial carbon pump, turbidity current carbon pump, bottom current carbon pump, and deep stratigraphic carbon pump, coupled with shallow-water and terrestrial systems, forming a “1+2” continuum transport system. Terrestrial system: Soil-Inland water organic matter continuous transport system; Shallow-water system: Land-Coast-Shallow shelf organic matter continuous transport system; Deep-water system: Shelf edge-Slope-Deep sea basin organic matter continuous transport system. FLI: Flux from land to inland waters; FIS: Burial in inland waters; FIA: Outgassing from inland waters; FIE: Flux from inland waters to estuaries; FEC: Flux from estuaries to continental shelf waters; FWS+FES: Carbon burial in tidal wetlands and estuaries; FAW-FEA: Carbon uptake by estuarine; FCS: Burial on continental shelves; FAC: Shelf water organic carbon uptake flux; FCO: Shelf-to-Deep sea organic carbon flux[10].) |
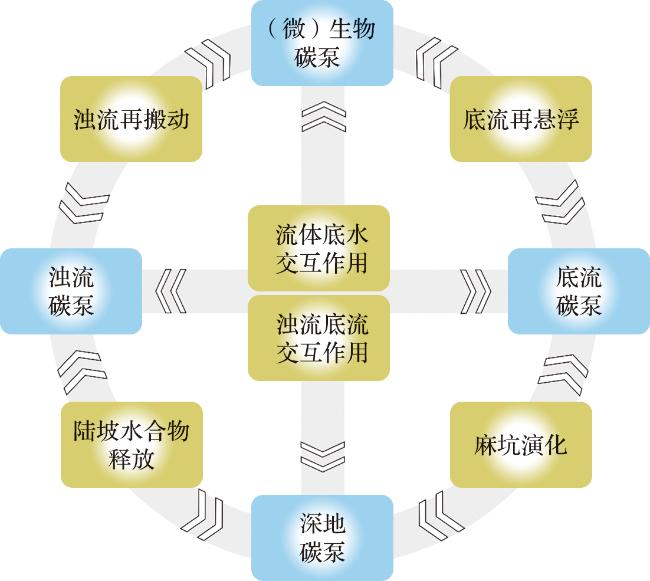
图2 深水体系有机碳泵交互耦合模式图(蓝色方框:具有鲜明特征与独立体系的碳泵端元; 黄色方框:不同碳泵端元间的典型交互方式或地质实例。) Fig.2 Schematic of the interacting-coupled model for the deep-sea organic carbon pump (Blue boxes: Represent carbon pump end-members with distinct characteristics and independent systems. Yellow boxes: Denote typical interaction mechanisms or geological examples between different carbon pump end-members.) |
2 深水浊流碳埋藏体系
表1 深水体系浊流碳泵分类方案Tab.1 A classification scheme for the turbidity current carbon pump in deep-water systems |
| 分类方案 | OC埋藏效率a/% | 发育频率/(次·a-1) | 全球典型实例 | |||
|---|---|---|---|---|---|---|
| 充填型浊流b | 冲刷型浊流c | |||||
| 峡谷体系 | 直接供给型 海底峡谷 | 河口相连型 | 60~100 | ≥10 | 1/(20~50) | 刚果峡谷、高屏峡谷 |
| 沿岸供给型 | ≥10 | 1/(100~1 000) | 蒙特雷峡谷、拉霍亚峡谷 | |||
| 陆架阻隔型 海底峡谷 | 跨越陆架型 | 60~90 | ≥10 | <1 | 孟加拉扇 | |
| 陆坡活跃型 | ≥1 | 1/100 | Whittard峡谷体系 | |||
| 陆坡休眠型 | 10 | 基本不发育 | 基本不发育 | 亚马孙峡谷体系 | ||
| 非峡谷体系 | 峡湾 | 80 | 100 | ≥1 | Bute峡湾 | |
| 大型海底滑坡 | 1/(2 000~30 000) | Storegga滑坡 | ||||
3 深水底流及深地作用碳埋藏体系
4 深水有机碳埋藏金字塔模型
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
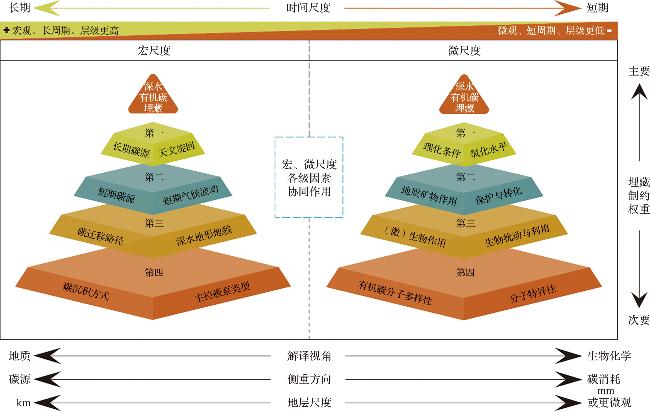
图3 深水沉积有机碳埋藏金字塔模型(本模型依据时间尺度、解译视角等方面将深水有机碳埋藏制约因素分属为两大类:宏尺度主要从地质-古海洋视角探讨碳源制约因素,微尺度主要从生物-化学视角探讨碳消耗、保存的制约因素,并依据各制约因素的相对重要性进行层级排序。) Fig.3 The pyramid model of organic carbon burial in deep-water sediments (This model classifies the constraints on deep-water organic carbon burial into two categories based on temporal scale and interpretive perspective: the macro-scale, which examines source constraints from a geological & paleoceanographic viewpoint, and the micro-scale, which focuses on consumption and preservation constraints from a biological & chemical viewpoint. These constraints are hierarchically ranked based on their relative importance.) |