
0 引言
传统蓝碳研究多聚焦于单一生态系统的内部碳循环过程(如植物光合固碳、沉积物埋藏),并已建立起相对清晰的碳收支模型[7-8]。然而,日益增多的证据表明,连续体内部的碳循环也存在显著的横向迁移。潮汐作为“天然泵”,驱动着凋落物、溶解有机碳(dissolved organic carbon,DOC)、颗粒有机碳(particulate organic carbon,POC)和溶解无机碳(dissolved inorganic carbon,DIC)在红树林与海草床之间进行高频、双向的交换[9-10]。这种跨生态系统的碳输送,不仅连接了相邻生境的碳循环,更从根本上改变了人们对单一系统碳源-汇格局的认知,进而导致当前的滨海蓝碳碳汇核算存在较大不确定性。目前对连续体碳循环的理解严重受限于碎片化的观测和单一过程的研究,对于温室气体交换通量、碳元素在水体-沉积物-生物多界面的传递路径以及沉积物碳库的真实来源与动态,均缺乏系统、同步的现场观测数据,且碳横向迁移的关键生物地球化学耦合机制仍有待揭示。
鉴于此,本文聚焦海草床-红树林连续体碳循环的最新研究成果,旨在从“纵向固存”与“横向迁移”两个核心维度,系统整合植物光合固碳、沉积物累积、水体碳转化及气体交换等关键过程,对其碳循环过程进行全景式梳理。同时,探讨当前研究的局限性与未来挑战,明晰关键科学问题,厘清连续体碳循环耦合机制的理论瓶颈;通过整合多技术方法,为突破观测与模拟的技术壁垒提供思路;以期为红树林和海草床修复项目的蓝碳潜力评估及其减缓气候变暖效益的量化提供理论依据。
1 海草床-红树林连续体及其碳循环
1.1 类型和特征
根据植被组成与地貌特征,可将海草床-红树林连续体划分为三种主要类型:边缘交错型、潮沟连通型以及镶嵌斑块型[11-12]。边缘交错型连续体是指红树林向海延伸至潮下带,与小型或匍匐型海草(如喜盐草属Halophila、二药草属Halodule)形成交错过渡带,如中国华南地区的秋茄-贝克喜盐草连续体[13];潮沟连通型连续体是指红树林通过潮沟系统与外围斑块状或带状海草床(如泰来草属Thalassia、丝粉草属Cymodocea)相连,有机质和颗粒碳通过潮汐泵作用在系统间迁移,如印度尼西亚Bintuni湾的红树林-海草系统[14];镶嵌斑块型连续体则是指在沉积动力较强的开阔岸段,红树林斑块与海草床呈镶嵌分布,碳交换以洋流扰动带来的颗粒碳再悬浮和沉积再分布为主,如澳大利亚Missionary湾的典型区域[10]。
海草床-红树林连续体的形成与维持依赖于红树林与海草床之间的正反馈机制。有证据表明,海草床可以在红树林前缘形成消浪带,保护红树林免受海浪冲击,促进红树林细颗粒物沉积与碳埋藏,使得沉积环境有利于红树林幼苗定植[6];而红树林则为海草床遮荫,减少光抑制,其凋落物输入在增加海草床沉积物沉积速率的同时,发达的根系结构还能稳定沉积物、降低侵蚀风险,从而保护海草床的生态完整性[5,15]。在具备高固碳能力的基础上,二者形成的连续体进一步依托潮汐作用,通过生态系统间协同互作实现碳循环耦合,提升滨海湿地的碳汇潜力[5]。海草床-红树林连续体碳循环过程表现出强烈的时空异质性,受潮汐周期、地貌类型、物种组成及人为干扰共同调控[16-17]。例如,大潮差区域的红树林凋落物输出通量显著高于小潮差环境,而海草床对外源碳的截留效率则随水体滞留时间增加而提升[12,18]。这种生态互惠机制显著增强了连续体对气候变化的适应能力,例如海平面上升背景下,红树林向陆迁移的同时,海草床可能向更深的潮下带扩展,从而形成新的碳汇热点[19]。
1.2 复杂的碳循环过程
2 海草床-红树林连续体的碳循环途径
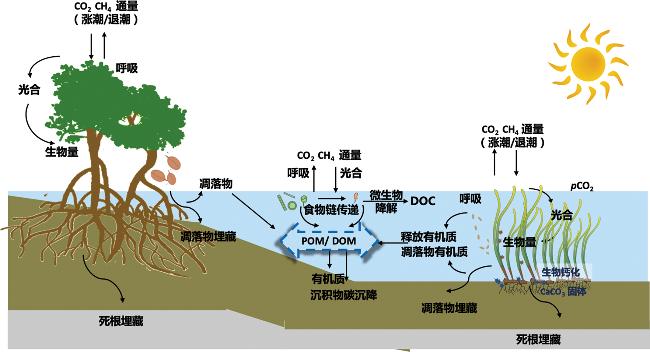
2.1 海草植物的光合固碳特征
植物光合作用是最主要的固碳过程。红树植物的光合固碳特征与陆地植物类似,其C3光合途径已被清晰地认知[7],而海草植物的光合途径则表现出多样化的特征。由于长期生活在潮下带,水下的弱光环境限制了海草植物的光合作用[28]以及根系生长,进一步影响地下碳的累积[29]。为适应海水中弱光、低CO2的环境,栖息在潮间带和潮下带的单子叶植物海草演化出了独特的光合适应策略。多数海草(如泰来草Thalassia hemprichii、喜盐草Halophila spinulosa、二药草Halodule uninervis、针叶草Syringodium isolifolium等)遵循C3光合途径。为尽可能利用水中无机碳,减少光呼吸带来的固碳损失,部分海草类群进化出了一系列区别于C3植物的碳循环机制:有研究表明,丝粉草Cymodocea nodosa、泰来草T. testudinum等部分物种,在低CO2环境胁迫下,能够激发一种基于CO2/HC${O}_{3}^{-}$泵和HC${O}_{3}^{-}$脱氢机制的C4型碳浓缩机制[30-32];另有喜盐草H. beccaeii等部分物种被认为属于C3-C4中间型[31](表1)。可见,海草植物的光合途径并不稳定,即使是同一个物种,在不同生境下,也可能具有迥异的光合代谢途径[33]。
表1 海草植物的主要光合途径Tab.1 Main photosynthetic pathways of seagrass |
| 光合途径 | 代表物种 | 主要分布区域 | 参考文献 | |
|---|---|---|---|---|
| C3途径 | 泰来草Thalassia hemprichii | 印度-西太平洋热带海域 | 文献[30-31] | |
| 棘状喜盐草Halophila spinulosa | 印度-西太平洋海域 | |||
| 单脉二药草Halodule uninervis | 热带及亚热带海域 | |||
| 针叶草Syringodium isoetifolium | 热带海域 | |||
| C4途径 | 结节丝粉草Cymodocea nodosa | 地中海、东大西洋沿岸 | 文献[30-32] | |
| 小丝粉草Thalassia testudinum | 加勒比海、墨西哥湾、西大西洋 | |||
| C3-C4中间型 | 贝克喜盐草Halophila beccarii | 印度-西太平洋热带及亚热带海域 | 文献[31] | |
| 诺氏鳗草Zostera noltii | 北大西洋、地中海、黑海及东亚温带海域 | |||
2.2 沉积物碳累积
除了植物碳循环外,海草床-红树林连续体纵向碳固存过程中,沉积物碳累积也是不可忽略的重要环节。全球滨海湿地的总初级生产力介于1.1~1.6 Pg·a-1之间[7-8,38],其中约16%的净初级生产产物储存在沉积物中[20]。连续体沉积物是红树林、海草床等滨海湿地储碳和固碳的主要区域,被认为是全球土壤碳储存的热点地区[39]。滨海湿地沉积物的有机碳具有两种不同来源,即内源性碳和外源性碳[40]。其中,内源性有机碳是沉积物纵向碳固存的主要来源。植物通过光合作用产生的有机碳,一部分通过根部钙化或者凋落物大量堆积的方式将碳直接固定在沉积物中[20-21],而沉积物的缺氧条件使有机碳的分解受到限制,从而实现了碳的长期储存[22];另一部分则通过呼吸作用、凋落物分解等过程,以CO2形式释放至水体或大气。凋落物分解过程中,部分碳将以POC的形式释放到水体中,进而再沉积,增加沉积物中的有机碳,提升系统整体的碳封存能力[22],并通过改变沉积物的物理化学性质,影响微生物的代谢活动和碳转化过程[17],从而调节系统的碳汇能力[5]。
2.3 水体碳循环
由于长期处于海水交替淹没的环境,海草床-红树林连续体的碳循环涉及多界面、多形式、多路径及多驱动因子的复杂过程。红树林产生的碳主要以溶解无机碳(DIC)和颗粒有机碳(POC)的形式输入邻近生态系统[7];相比之下,海草床系统不仅输出DIC和POC,其水体中还赋存大量溶解有机碳(DOC)和颗粒无机碳(PIC),参与碳循环[45-46]。多种过程都会对海草床-红树林连续体的CO2源-汇格局产生不同影响:例如水体中海草以生成碳酸钙的形式释放CO2,使得水体总碱度(TA)降低,从而促进CO2向大气释放,而碳酸钙的溶解过程则产生相反效应[47];沉积物中的微生物代谢过程是水体TA的主要来源,其通过沉积物-水界面的碳交换影响水体的pH与CO2分压(pCO2)[48-49]。此外,自养生物的初级生产、有机物矿化作用等多种因素的耦合会进一步影响水体pCO2,最终共同塑造海草床-红树林连续体的CO2源-汇格局。
在淹水状态下,海草和红树植物经光合作用将溶解无机碳转化为有机碳并贮存在生物量中,其中部分有机碳以DOC形式侧向渗出至水体[50-52],而植物碎屑及浮游生物残骸则以POC形态悬浮于水体[53]。在POC的再矿化过程中,DOC作为中间产物被二次释放,二者共同构成群落净生产和跨界面碳交换的活性组分[54]。随后,DOC在微生物降解中经历两个主要去向:一部分经呼吸作用转化为CO2释放到大气中[55],另一部分则转化为难降解、难溶解的惰性溶解有机碳(refractory dissolved organic carbon,RDOC)。RDOC构成了海洋DOC的主体[56],其化学惰性赋予其百年至千年尺度的存留潜力,成为蓝碳长期封存的关键形态。然而,针对海草床-红树林连续体RDOC的存量与生成通量,迄今仍缺少系统的现场监测与定量评估。
2.4 气体交换
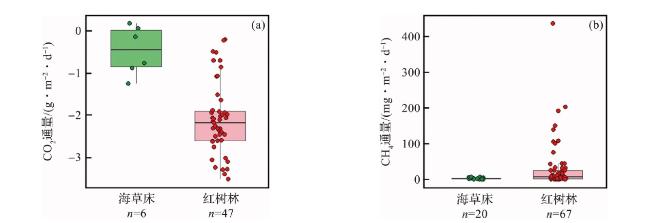
海草床-红树林连续体的碳循环主要依赖于光合作用、呼吸作用、分解作用等过程,实现生态系统与大气之间的碳交换[26]。综合大量研究发现,红树林贡献的CO2吸收量占全球沿海植被CO2吸收总量的60%以上,全球海草的CO2吸收量(192 Tg·a-1)约为盐沼的4倍 ;同时,红树林也是沿海植被CH4排放量的主要来源,排放通量高达30.6±8.3 mg·m-2·d-1,而海草的CH4排放通量(1.8±0.4 mg·m-2·d-1)相对较小,约为红树林的6%(图2)。然而,受限于水下观测困难,完整潮汐周期内海草床和红树林的碳通量过程并不十分清晰。在涨潮时,植物浸淹于水下,在弱光和低CO2环境中,连续体主要通过水体中的CO2交换实现碳循环[29];在退潮期间,潮间带植物暴露在大气中持续数小时之久,同时伴有干燥和辐射增加等条件,此时与大气间的CO2交换占碳循环主导地位。因此,受潮汐、光照、温度、藻类等多重环境与生物因子共同调控,海草床、红树林的CO2交换呈现日尺度与季节尺度的复杂变化[16,57]。在海草床-红树林连续体的厌氧沉积物中,丰富的有机质为产甲烷古菌提供了活跃的底物,驱动甲基营养型等多种代谢途径生成甲烷,成为厌氧条件下碳转化与输出的关键路径。由此产生的甲烷排放会显著抵消系统的碳汇功能,导致对净气候效益的高估:研究表明,红树林沉积物CH4排放可抵消其固碳效益的27.5%[58];热带海草床的CH4排放率通常高于温带区域,全球海草床的CH4排放可在20年的时间尺度上抵消其碳埋藏效益的8.6%[59]。另外,前人在研究滨海湿地碳循环的过程中往往聚焦于独立生态系统中植物或沉积物的碳交换过程,而忽略了由于潮汐作用引起的CO2与CH4交换[60];受海草长期浸没造成的观测条件限制,针对海草床的气体交换研究仍显不足,相关数据不足红树林的1/3(图2)。因此,海草床-红树林连续体在不同潮汐状态下的CO2通量格局仍有待解析,忽略潮汐对碳循环的影响将导致连续体碳汇潜力的估算产生误差。
3 海草床-红树林连续体横向碳迁移过程、
通量与耦合机制 滨海湿地中的海草床-红树林连续体具有海陆双重特征,其有机碳的来源、转化和归宿难以用单一的生态系统边界进行划分。横向碳迁移是指从红树林、海草床输出至邻近生态系统的碳交换过程,既包括地表产生的POC通过潮汐作用发生的迁移,也涵盖地下间隙水驱动的横向输出。近十年来,稳定同位素技术(δ13C、δ15N、Δ14C)、放射性核素年代学(210Pb、137Cs)以及高分辨率水动力-生物地球化学耦合模型(Delft3D-BIO, FVCOM-ICM)被广泛应用于海草床-红树林连续体碳迁移研究。结果表明:连续体内部的物质与能量循环并非封闭系统,而是在潮汐、风浪以及生物扰动等多重驱动下,进行着高频、双向的横向交换[9,11]。这种跨系统的碳耦合显著提升了水-沉积物界面的碳汇效率,并可能因海平面上升和人类活动干扰而呈现非线性响应[69-70]。
3.1 海草床碳迁移:外源碳的“汇”与颗粒碳的“源”
海草床作为典型的沉水植被生态系统,其碳固持能力不仅取决于海草自身的净初级生产(net primary production,NPP),更受到大量外源有机碳输入的强烈调控。综合全球114处海草床沉积物的同位素质量平衡分析结果可知,海草床表层0~50 cm沉积物中的有机碳平均仅有约52%±11%直接来源于海草初级生产,其余约48%则来自外源输入,包括红树林凋落物、陆源土壤有机质、河流输送的浮游植物残体以及近岸浮游动物粪球颗粒等[2,20]。在印度尼西亚Bintuni湾的研究中,三端元混合模型计算显示,红树林凋落物对海草床沉积物有机碳的贡献可达30%~45%,且贡献量随潮差增大呈线性上升趋势[14]。
此外,海草床水体是连续体中POC向外输出的主要节点。SANTOS等[71]基于全球78个海草床24 小时围隔通量观测与遥感反演结果,估算出海草床POC的年输出通量为87±21 Tg·a-1,与同期沉积物有机碳年埋藏通量(83±19 Tg·a-1)基本相当,表明海草床在空间格局上具有“近源-远汇”的双重功能。值得注意的是,海草床输出POC的δ13C值介于-24.5‰~-18.9‰之间,明显受红树林C3植物信号影响,进一步佐证了连续体间的横向碳输送[18]。最新的浮标-滑翔机联合观测显示,海草床向邻近水体释放的DOC呈现显著的日变化特征:白昼以光依赖的光合作用为主(0.8~1.4 mmol·m-2·h-1),夜间则随着呼吸作用与微生物再矿化同步增强(2.1~3.6 mmol·m-2·h-1),其中约15%~30%的DOC可进一步转化为RDOC,并随潮流向外海输送[54]。
3.2 红树林碳迁移:高生产-高输出的碳泵
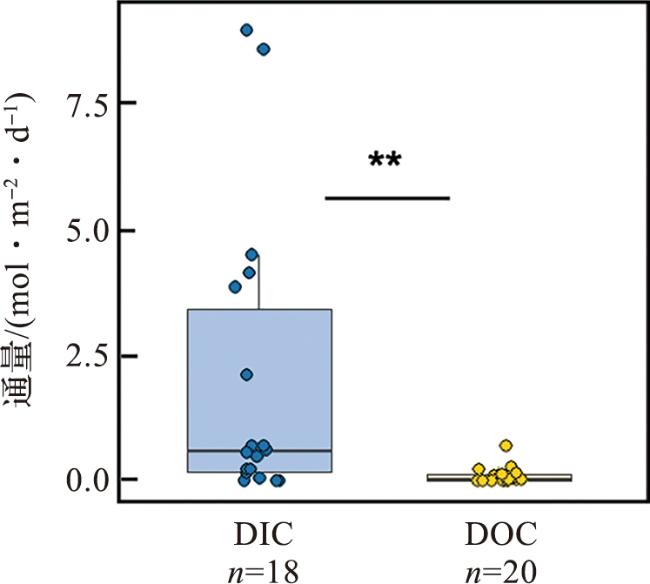
红树林虽被誉为滨海湿地最大的碳库[72],但其固定的碳有相当一部分通过潮汐运动、孔隙水排放以及凋落物掉落等过程输出到邻近的水体中[73]。在潮汐作用下,红树林以POC等形式向近岸海水输出的碳量,可占其沉积物固碳总量的50%以上[72,74]。例如,在澳大利亚Missionary Bay,涡度相关-潮位同步观测系统的数据显示,红树林向海草床输出的POC瞬时通量可达44±9 g·m-2·d-1,相当于同期红树林净生态系统生产(net ecosystem production,NEP)的53%[10]。在亚马孙河口,红树林输出的DOC浓度高达6.3±1.7 mmol·L-1,其浓度沿盐度梯度呈指数衰减,半衰减距离约为8.5 km,这些DOC直接支撑了下游海草床的异养呼吸过程[75]。此外,红树林根系呼吸和硫酸盐还原过程释放的DIC(平均浓度为3.9 mmol·L-1)可通过密度流沿潮沟底部向海草床输送,形成“暗碳”通道[46]。若忽视这些横向碳输出,可能导致区域尺度上红树林碳汇估算偏高30%~110%[76]。大量研究表明,DIC是红树林与邻近生态系统碳交换的主要形态[77-79],红树林通过地下水交换输出的DIC和DOC通量分别为1.99±0.67 mol·m-2·d-1(n=18)和0.12±0.04 mol·m-2·d-1(n=20)(图3)。全球红树林DIC输出通量为(1.03~2.23)×1013mol·a-1,约占全球河流向海洋输送DIC总量的46%[80]。全球红树林DIC和DOC通量远大于红树林碳埋藏总量(3.42×1012 mol·a-1)[81]。因此,横向输出的DIC是红树林生态系统不可忽视的碳输出(图4)。最新一项涵盖东南亚12个河口区的研究指出,将横向输出(POC+DOC+DIC)纳入碳收支后,红树林净生态系统碳平衡(net ecosystem carbon balance,NECB)平均下调46%[17]。由于红树林与其他蓝碳生态系统间的碳交换通常基于碳储量与总生产力之间的差值间接估算,而现有研究缺乏对连续体间碳交换过程的解析,这可能导致红树林碳汇潜力的高估超过一半[76]。

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
3.3 海草床-红树林连续体耦合碳循环
由于海草具有密集、柔韧的茎叶结构,红树则发育复杂的地上根系,二者共同构成一张“生物过滤网”,在涨落潮过程中截获大量悬浮有机质并促进其沉降。研究认为,海草床和红树林之间互为碳源:红树林超过一半的固定碳通过潮汐作用进入海草床及邻近生态系统,而海草床亦有约1/4的净初级生产力以DOC或POC的形式输出至红树林(图4)。稳定同位素混合模型与脂肪酸生物标志物分析结果均表明,在华南淇澳岛连续体,海草床表层0~5 cm的沉积物中,有83%±6%的有机碳来源于红树林凋落物;而在红树林表层沉积物中,亦有21%±4%的碳呈现海草典型的δ13C特征(-10.2‰~-6.8‰),印证了双向输送的普遍性[15]。在澳大利亚东北部,气候变暖促进植物生长的同时,也显著提高了沉积速率[40];多核素(210Pb、137Cs)沉积记录显示,过去150年来红树林-海草床连续体表面沉积速率提高了2.3倍,其与海平面上升引发的潮沟溢流事件更是频繁同步,说明横向碳通量对气候变化可能具有正反馈效应[19]。除CO2交换外,海草床-红树林连续体兼具红树林的高CH4生产潜力与海草床的高CH4生成速率,可能形成独特而活跃的CH4产生与排放环境,成为CH4排放的潜在热点。忽略这种协同或叠加效应,将对连续体净全球增温潜势的评估造成显著偏差。
海草床-红树林连续体通过潮汐、颗粒有机碳再悬浮与生物过滤等多种机制耦合,实现了碳的横向迁移与再分配。明确连续体内部的“横向迁移—再悬浮—再埋藏”完整路径,建立时空分辨率更高的潮动力-生物地球化学耦合模型,是准确评估滨海蓝碳汇及其对未来气候变化和人类活动响应的关键。
4 存在问题与展望
海草床-红树林连续体的碳循环研究虽取得了显著进展,但在碳汇功能的定量评估与预测方面仍面临严峻挑战。连续体间的碳交换受到极为复杂的物理和生物过程机制共同调控,导致厘清其碳循环过程存在壁垒。未来研究需重点关注以下五个关键方向,以深化理论认知并提升其对气候减缓行动的支撑价值。
4.1 连续体多界面碳交换的耦合机制亟待厘清
当前对海草床-红树林连续体碳循环的理解多局限于单一生态系统或单一过程,对于潮汐驱动下“红树林-水体-海草床-沉积物-大气”等多界面间的碳交换耦合机制仍认识不足。例如,凋落物分解、根系分泌物释放、沉积物-水界面扩散、水体垂向混合以及海-气交换等过程,如何在小时至季节尺度上协同响应潮汐动态与气候事件,尚不明确。理论突破的关键在于揭示这些多界面耦合与反馈机制,从而系统理解连续体作为整体,其碳汇功能的稳定性与韧性。在研究手段上,亟需发展能够同步监测多界面碳通量的原位观测系统,并结合稳定同位素、微生物功能基因测序等技术,追溯碳的迁移路径与转化过程。
4.2 气体交换通量观测的时空局限
目前,全球范围内对海草床进行长期、连续大气碳通量观测(如采用涡度相关法)的站点屈指可数,主要集中在澳大利亚、地中海和美国的少数几个温带及热带海域的典型区域(如澳大利亚的Cleveland Bay、西班牙的Alfacs Bay)。在我国南方典型连续体中,针对小型海草(如贝克喜盐草Halophila beccarii)及其与红树林交错带在潮间带尺度上的气体交换过程,尚缺乏精确量化研究,导致无法准确评估在不同潮汐淹没状态(浸淹 vs.暴露)、昼夜及季节尺度上的海草床碳源-汇动态,也难以厘清其在连续体碳收支中的净贡献。
4.3 多形态碳元素的跨界面传递路径不清
连续体内碳的横向迁移以DOC、POC和DIC为主要载体。然而,现有研究多依赖于离散的水样采集和实验室分析,无法捕捉潮汐驱动下的高频、动态变化;对于红树林凋落物在潮沟中的破碎与淋溶过程,以及退潮期间海草床是否向大气释放源自红树林的碳等关键迁移路径,仍缺乏连续的现场实测数据支撑;同时,微生物介导的DOC转化与RDOC生成过程几乎未被现场观测覆盖。上述不足使得稳定同位素等溯源技术的应用因缺少端元信息和过程数据而受限。
4.4 沉积物碳累积与来源的异质性认知不足
尽管沉积物柱状样采集法被广泛用于评估碳埋藏,但在海草床-红树林连续体中,沉积环境具有高度的空间异质性。一方面,水动力分选、生物扰动(如蟹类、多毛类)会导致碳的再悬浮和再分布;另一方面,传统210Pb测年法在潮间带环境中易受沉积物再悬浮与混合作用影响,使得通过传统210Pb测年法获得的单点沉积物柱状样沉积速率难以代表整个连续体的平均状况,从而造成碳埋藏速率的估算误差。同时,关于沉积物碳来源(内源 vs.外源)的贡献率,现有研究多基于有限参数的混合模型进行估算,缺乏对不同潮汐条件、不同季节下外源碳输入通量和归宿的直接观测。尽管稳定同位素(δ13C、δ15N)与生物标志物(如木质素酚类)可用于区分红树林源碳与海草源碳,但外源藻类有机碳与陆源有机碳的贡献往往难以剔除,导致碳源解析结果存在较大不确定性。
4.5 横向碳通量的定量监测与模型模拟能力不足
横向碳迁移是连续体碳汇功能的核心环节,但如何对其进行量化仍是巨大挑战。忽略高频、双向的横向碳交换可能导致对整个系统净碳平衡的高估或误判(如忽略红树林通过DIC形式向海草床及外海的“暗碳”输出)。理论上的关键进展将依赖于构建能够精确量化横向碳通量的方法论体系。同时,研究手段也亟需创新,包括布设高分辨率断面监测网络、应用水面无人机和遥感技术追踪有色溶解有机物(colored dissolved organic matter,CDOM)等示踪物,并重点开发耦合水动力-生物地球化学过程的高分辨率数值模型(如FVCOM-ICM),以模拟在不同海平面上升和人类活动情景下的碳通量变化。
此外,当前的海草床或红树林修复项目,大多基于单一的生态系统服务目标(如防灾减灾、生物多样性),较少从整体提升连续体碳汇功能的角度进行系统性规划。如何通过优化空间配置(如红树林与海草床的宽度、连通性)、选择关键物种组合以及实施水文恢复措施,以增强碳的捕获与封存效率,是亟待解决的应用科学问题。未来应加强“修复生态学”与“生物地球化学”的交叉研究,通过设置控制实验与长期监测,识别碳汇效率最高的连续体构型,发展“增汇型修复”范式,从而为基于自然的气候变化解决方案提供精准的工程设计与管理实践指导。