
0 引言
自工业革命以来,化石燃料的大量使用以及人类活动的增加导致大气CO2含量不断升高,全球大气CO2平均体积分数已从工业化前(1750年前)的约278×10-6增加至2023年的420×10-6,并且最近10年的增速达到了2.4×10-6/a[1]。由CO2排放增加引发的温室效应、海洋酸化、海平面上升等全球环境问题日趋严重。河口连接了陆地和海洋,既是物质由陆地向海洋输送的主要渠道和天然、高效的过滤器,也是生物地球化学反应池。在河口区,由于水体携带大量颗粒有机物,浊度通常比较高,抑制了浮游植物的光合作用,在颗粒有机物的氧化、降解过程中,消耗氧气的同时产生了大量CO2释放到大气中,使河口区成为CO2的源[2]。随着颗粒有机物的沉降与降解,水体浊度降低,浮游植物吸收CO2进行光合作用,使河口区转换为CO2的汇[3]。受外源物质输入和内部活跃的生物地球化学反应的共同影响,河口区水体理化参数的空间异质性高,这导致对该区域海-气CO2通量估算的不确定度较高。因此,增加不同类型河口的观测数据是目前河口碳循环研究的现实需求。
红树林主要分布在热带和亚热带海岸的潮间带,全球总面积约为1.4×105km2[4],仅占陆架边缘海面积的2%,但却贡献了占全球大约5%的净初级生产力、12%的生态系统呼吸以及占亚热带和热带海域大陆架约30%的碳埋藏,其溶解无机碳(DIC)的输出占所有流入低纬度沿海水域河流DIC通量的1/3[5]。因初级生产力和土壤碳埋藏速率高,红树林的固碳能力非常强,全球红树林碳储量约为4.03 Pg(1 Pg=1015 g),在赤道附近、10°N—20°N和20°N—30°N,碳储量分别为2.72 Pg、1.00 Pg和0.29 Pg[6]。全球红树林沉积物碳埋藏速率约为38.3 Tg·a-1[4]。红树林通过潮汐泵向周边海域输送不同碳组分,其中DIC为主要形态(86 Tg·a-1),颗粒有机碳(POC)次之(28 Tg·a-1),溶解有机碳(DOC)占比最少(5 Tg·a-1)[7]。虽然不同碳组分的迁移、传输过程存在差异,但其最终都汇集于邻近海域,通过呼吸作用、光合作用等生物地球化学过程影响着近海CO2的源汇格局。一般认为,红树林生态系统为大气CO2的汇,其周边潮沟为大气CO2的强源,但也有研究发现,由于碱度(TA)的横向输运与积累,红树林生态系统的邻近海域会成为大气CO2的汇[7]。因此,当有潮沟水体和河流同时输入时,会使得河口区的CO2源汇格局变得复杂,这在干湿季分明的热带河口特别明显。
本研究通过现场调查与室内分析,探究了东寨港干湿季CO2分压(pCO2)分布、影响因素以及CO2通量,以期解析热带红树林河口CO2源汇格局,也为当地生态环境综合治理提供理论依据和技术支撑。
1 研究区域
2 数据获取和处理
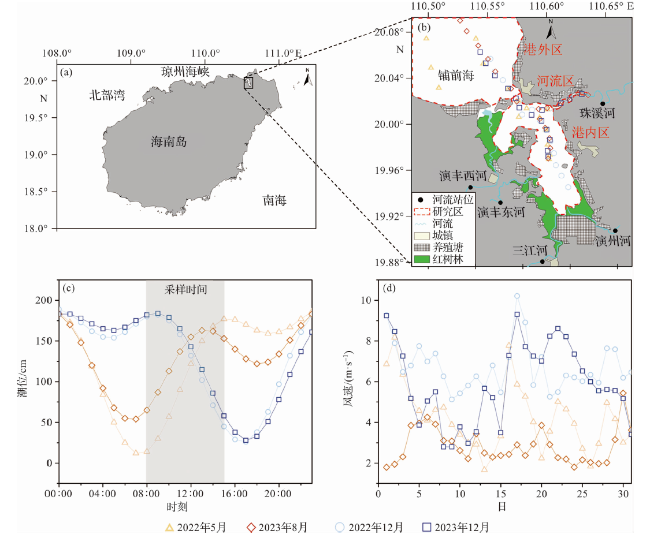
将研究区分为河流区(珠溪河)、东寨港内、东寨港外(铺前海)三个子区域(图1b),分别于干季(2022-12-23、2023-12-27)、湿季(2022-05-20、2023-08-20)开展4个航次调查,采集表层水体(0.5 m)用于理化指标和碳酸盐参数的测定,采样时间在8:00~15:00,其中2022年5月航次未采集珠溪河数据。在5条河流的上游(不受潮汐影响的区段),各设置一个站位采集上述样品(图1b),采集时间为航次的前一天。干季时河流的径流量较小,河口整体盐度较高,采样在退潮时进行(图1c);湿季时河流径流量较大,河口整体盐度较低,采样在涨潮时进行。船载采样当天的潮位数据来自国家海洋科学数据中心(https://nmdis.org.cn),采样当月的风速数据来自欧洲中期天气预报中心(https://www.ecmwf.int)。
使用HI98192测定仪(HANNA)现场测量水样的温度和盐度。DIC、TA、pH、溶解氧(DO)的测定参照全球碳通量联合研究计划(JGOFS)的推荐方法[12],DIC的测定采用非分散红外吸收法(AS-C3, Apollo SciTech);TA的测定采用Gran滴定法(AS-ALK2, Apollo SciTech),用海水无机碳标准品(美国Scripps海洋研究所)进行校正,精度为2 μmol·kg-1;pH的测定采用玻璃电极法,精度为0.01;DO的测定使用Winkler光度法(V-1200, MAPADA),并进行了浊度校正,精度为1 μmol·kg-1。pH与DO的测定在采样后的48 h内完成。
饱和溶解氧通过现场测定的温度、盐度计算得到,其与实测溶解氧的差值称作表观耗氧量(apparent oxygen utilization,AOU)。实测溶解氧与饱和溶解氧的比值为溶解氧饱和度。根据DIC、TA、温度、盐度计算得到表层水体的pCO2,水体pCO2与大气pCO2(421 μatm,来自其他船载走航观测结果)的比值为pCO2饱和度。
单位面积、单位时间的海-气CO2通量(F)通过以下公式计算:
F=k·α·(pwaterCO2-pairCO2)
k=(0.314· -0.436·u10+3.99)·(Sc/600)-0.5
式中:α是一定温度、盐度下水体中CO2的溶解度;Sc为施密特数;k为气体交换速率(单位:cm·h-1),为特定条件下风速的幂函数,一般采用海平面10 m处的风速值(u10)。本文结合研究区域特征,对k值的计算采用JIANG等[15]的方法。
区域海-气CO2通量(FA)采用以下公式进行计算:
式中:An为不同区域的面积,Fn为单位面积、单位时间海-气CO2通量的平均值。
3 结果
3.1 东寨港上游河流水体理化特征
河流携带大量淡水和营养物质进入河口,影响了河口水体中生源要素的循环。航次期间, 5条河流水体的理化参数存在干湿季差异(表1)。水温干季平均值(18.1~19.0 ℃)低于湿季(27.9~30.0 ℃);盐度平均值在干湿季无明显差异;pH干季平均值(6.78~6.94)高于湿季(6.26~6.58);除珠溪河外,其余4条河流的DIC与TA平均值均为干季高于湿季;DO干季平均值比湿季高42~83 μmol·kg-1,且所有河流均处于溶解氧的亏损状态(DO饱和度:59%~90%),AOU为29~95 μmol·kg-1;除演丰西河外,其余4条河流的pCO2平均值均为干季低于湿季。调查期间,河流pCO2平均值是大气pCO2的8.5倍,表现为大气CO2的强源。
表1 东寨港周边主要河流水体各理化参数的平均值Tab.1 The average values of various physical and chemical parameters of major rivers around Dongzhai Harbor |
| 河流 | 季节 | 温度/℃ | 盐度 | pH | DIC /(μmol·kg-1) | TA /(μmol·kg-1) | DO /(μmol·kg-1) | DO饱和度/% | AOU /(μmol·kg-1) | pCO2 /μatm |
|---|---|---|---|---|---|---|---|---|---|---|
| 演丰西河 | 干季 | 18.1 | 0.10 | 6.89 | 1 126 | 1 048 | 234 | 79 | 62 | 2 941 |
| 湿季 | 27.9 | 0.03 | 6.58 | 877 | 757 | 192 | 78 | 54 | 2 761 | |
| 演丰东河 | 干季 | 18.3 | 0.09 | 6.81 | 854 | 730 | 224 | 76 | 71 | 3 657 |
| 湿季 | 29.0 | 0.02 | 6.34 | 750 | 600 | 168 | 68 | 74 | 4 318 | |
| 三江河 | 干季 | 18.7 | 0.06 | 6.85 | 749 | 618 | 259 | 90 | 29 | 3 501 |
| 湿季 | 29.4 | 0.01 | 6.34 | 689 | 549 | 183 | 75 | 57 | 4 511 | |
| 演州河 | 干季 | 19.0 | 0.06 | 6.78 | 672 | 535 | 226 | 77 | 69 | 2 990 |
| 湿季 | 30.0 | 0.04 | 6.27 | 640 | 520 | 143 | 59 | 95 | 4 939 | |
| 珠溪河 | 干季 | 18.1 | 0.10 | 6.94 | 661 | 544 | 224 | 75 | 76 | 2 113 |
| 湿季 | 28.8 | 0.10 | 6.26 | 780 | 693 | 178 | 73 | 64 | 3 900 |
3.2 东寨港及周边水域水体理化特征
图2为东寨港及周边水域4个航次的调查结果,各理化参数在港内、港外和珠溪河三个子区域的表现各不相同。温度的空间变化幅度小,在港外、港内和珠溪河的分布相对均匀。港内外水体的盐度维持在较高水平(28.93±3.25),由港外向港内逐渐递减,珠溪河的盐度显著低于港外和港内,变化明显。干季DO的空间差异较大,湿季差异不明显。pH和盐度呈现相同的趋势,由港外向港内逐渐递减,珠溪河最低。水体pCO2的空间分布大体与盐度、pH相反,在整个研究区域均高于大气pCO2(421 μatm),即研究区是大气CO2的源。
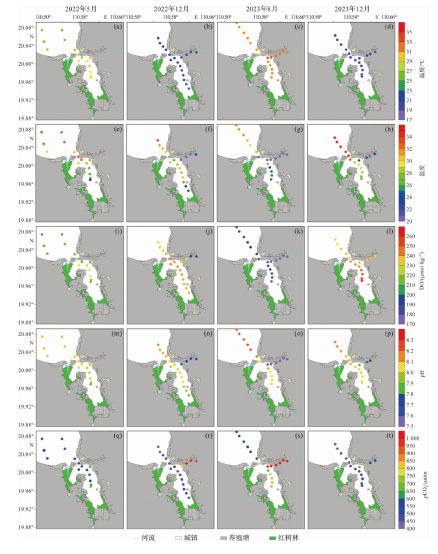
东寨港及周边水域水体理化参数存在明显的干湿季差异。珠溪河,干季盐度平均值(18.92±3.77)、pH平均值(7.74±0.08)、DO平均值(216±16 μmol·kg-1)、DO饱和度平均值(84%±6%)高于湿季,干季温度平均值(19.0±0.5 ℃)、pCO2平均值(760±170 μatm)低于湿季。
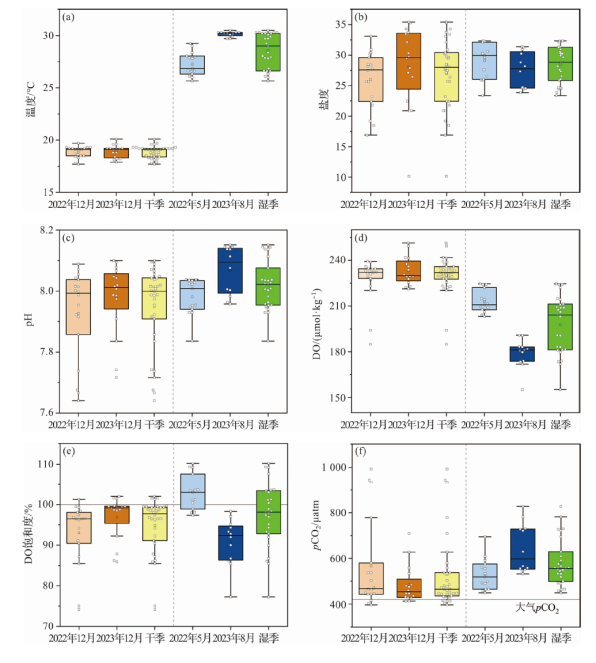
东寨港内、外水体理化参数的干湿季差异见图3。航次期间,8月份水温平均值(30.1±0.2 ℃)最高,5月份(27.3±1.1 ℃)次之,12月份较低且两个航次差别不大。干湿季的平均盐度差别不大,但干季的变幅大于湿季。干湿季节pH平均值差别不大。干季DO高于湿季,但是干湿季的DO饱和度平均值接近。pCO2的季节变化与DO相反,湿季高于干季,2023年8月最高,2023年12月最低。
3.3 影响东寨港水体pCO2的因素
3.3.1 温度
温度通过影响CO2溶解度和碳酸盐体系解离过程来调控水体pCO2,通常温度每升高1 ℃,海水pCO2增加4%~5%[13]。东寨港的海水源自南海北部陆架表层水,已有研究显示,该水团年平均温度为26.0 ℃,年均pCO2为370±20 μatm,pCO2随温度的变化曲线基本分布在350e0.042 3(t-26)和390e0.042 3(t-26)两条参考线之间(图4a黑色虚线)[16],表明温度是南海北部陆架表层水pCO2的主要调控因素。由于存在多种外源输入和内部活跃的生物地球化学过程,影响东寨港水体水pCO2分布的因素较南海北部陆架表层水更为复杂。干季、湿季pCO2分别为396~993 μatm和450~827 μatm,均高于南海北部陆架表层水(图4a)。干季温度变幅在2.0~2.2 ℃,其引起pCO2的变化量(pCO2与NpCO2的差值)为22~27 μatm,占pCO2变幅(296~597 μatm)的3.6%~9.1%;湿季温度变幅为2.4~3.6 ℃,其所引起pCO2的变化量为44~63 μatm,占pCO2变幅(245~295 μatm)的14.8%~25.6%。该结果说明温度对东寨港pCO2有影响,但不是主要的影响因素。
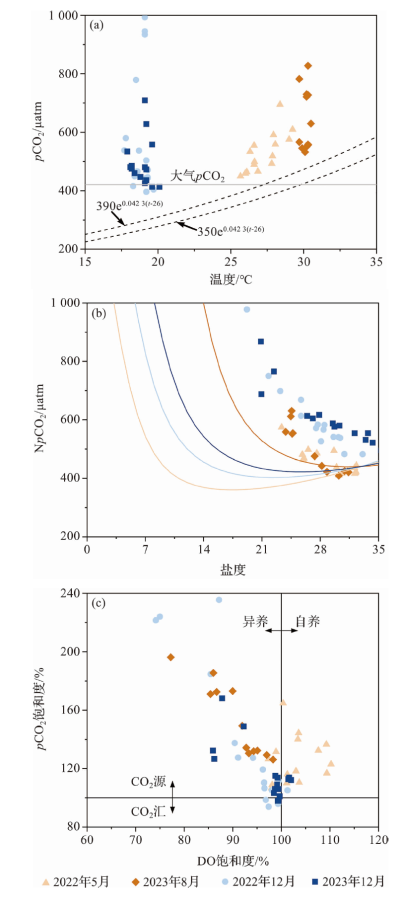
图4 温度与pCO2的分布(a)、盐度与NpCO2的分布(b)以及DO饱和度与pCO2饱和度的分布(c)(图b中曲线为仅发生物理混合时NpCO2随盐度的分布。) Fig.4 Distributions of temperature and pCO2 (a), salinity and NpCO2 (b), and dissolved oxygen saturation and pCO2 saturation (c) (Curves in Fig.b show the distribution of NpCO2 with salinity when only physical mixing occurs.) |
3.3.2 物理混合
3.3.3 生物过程
由于生物的光合作用与呼吸作用会影响水体的DO与pCO2,因此可以通过DO饱和度与pCO2饱和度的变化来探讨水体中光合作用/呼吸作用的强弱。当光合作用大于呼吸作用时,水体中DO饱和度大于100%(即自养),而当呼吸作用大于光合作用时,水体中pCO2饱和度将大于100%(即CO2源)[18]。图4c显示,大部分数据都分布在第二象限,即DO饱和度降低的同时pCO2饱和度升高, 两者呈负相关关系,说明呼吸作用主控pCO2的变化。2022年12月与2023年8月在珠溪河下游站位出现了DO饱和度低值(<80%)、pCO2饱和度高值(>190%)的现象,说明下游水体的呼吸作用强度大。2022年5月的大部分数据分布在第一象限,即水体处于自养和CO2源的状态,说明在DO过饱和的情况下,水体仍然向大气释放CO2。推测该航次调查期间,存在着明显的CO2外源输入,例如河流输入、沉积物间隙水向上扩散、碳酸钙生成等,并且CO2输入量超过了光合作用所能吸收的CO2量。在墨西哥湾、长江口等地均曾出现过类似现象[19-20]。
3.4 海-气CO2通量
3.4.1 东寨港及周边水域CO2通量的时空变化
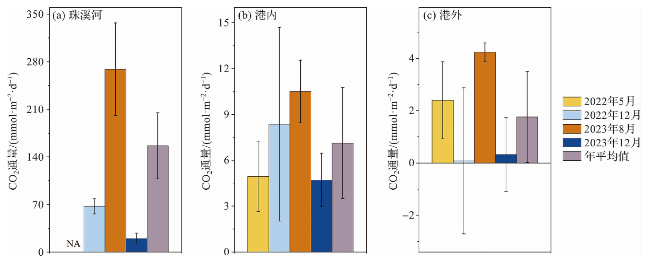
整个研究区域均向大气释放CO2,但各子区域的CO2通量差异大,数值从珠溪河(156.3±48.6 mmol·m-2·d-1)向港内(7.1±3.6 mmol·m-2·d-1)、港外(1.8±1.7 mmol·m-2·d-1)递减(图5)。以珠溪河作为河流的代表,通过面积权重法求算海-气CO2通量,整个研究区的区域年平均CO2通量约为6.1±6.3 mmol·m-2·d-1,其中河流、港内与港外水体的贡献量分别为36%、47%和17%。从该结果可知,虽然河流的面积(1.33 km2)远小于港内(38.41 km2)和港外(55.21 km2)水域,但其对研究区CO2通量的贡献不容小觑。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
航次期间,各区域CO2通量多表现为湿季大于干季,其中,河流的湿季平均值(269.0±68.1 mmol·m-2·d-1)远高于干季(43.5±9.7 mmol·m-2·d-1);港内的湿季平均值(7.7±2.2 mmol·m-2·d-1)略高于干季(6.5±4.6 mmol·m-2·d-1);港外干季整体上处于与大气接近平衡的状态(0.2±2.2 mmol·m-2·d-1),而湿季则是CO2的弱源(3.3±1.1 mmol·m-2·d-1)。整个研究区湿季平均值(8.8±8.2 mmol·m-2·d-1)是干季(3.4±3.6 mmol·m-2·d-1)的2.6倍。
从公式(1)可知,海-气CO2通量主要受到气体交换速率k (风速的经验公式)与海-气CO2分压差的影响。航次期间,湿季的平均风速(3.4 m·s-1)约为干季(6.2 m·s-1)的1/2(图1d),而湿季的海-气CO2分压差远大于干季,其中河流、港内、港外水域的湿季CO2分压差分别达到了干季的17.5、4.0和33.0倍。可见,相比风速,海-气CO2分压差对东寨港CO2通量的影响更大。
3.4.2 东寨港CO2通量与全球红树林河口比较
为探究东寨港CO2通量在全球红树林河口中的水平,汇总了全球40个红树林伴生河口表层水pCO2变化范围与CO2通量的数据[21⇓-23](表2),发现红树林河口的CO2通量均大于0,其中通量值在0~10 mmol·m-2·d-1和10~50 mmol·m-2·d-1的分别占22%和29%,其余的红树林河口(n=21,49%)均是CO2的强源(>50 mmol·m-2·d-1)。与其他红树林河口相比,东寨港的CO2通量(6.1 mmol·m-2·d-1)处于较低水平,与同纬度的Sundarbans、Mooriganga河口相当,远低于同纬度的Dhamra河口与Itacuraca河口。CO2通量存在干湿季差异是红树林河口的普遍特征,例如澳大利亚Fitzroy 河口,湿季(215.2 mmol·m-2·d-1)是干季(73.1 mmol·m-2·d-1)的2.9倍;亚马逊Mojuim River河口,干季(658.8 mmol·m-2·d-1)是湿季(230.8 mmol·m-2·d-1)的2.9倍[24]。东寨港CO2通量较低,可能的原因之一是计算时纳入了低通量的港外区域,若只考虑港内与河流水域,则数值增大近一倍(12.1 mmol·m-2·d-1)。另外,CO2通量较低可能与气体交换速率k仅考虑风速相关。已有研究表明,除了风速以外,潮汐、流速、水深和水域开阔度也是影响近海浅水区气体交换速率的重要因素[25-26]。
表2 东寨港与其他红树林河口[21⇓-23]的CO2通量比较Tab.2 Comparison of CO2fluxes among Dongzhai Harbor and other mangrove estuaries[21⇓-23] |
| 国家/地区 | 河口 | 水体pCO2 /μatm | CO2通量 /mmol·m-2·d-1 |
|---|---|---|---|
| 安达曼群岛 | Kalighat | 1 574~7 888 | 70.8 |
| 安达曼群岛 | Wright Myo | 1 246~7 703 | 61.1 |
| 澳大利亚 | Moreton Bay | 9 513~27 188 | 201.6 |
| 澳大利亚 | Constant Creek Estuary | 449~1 632 | 21.5~50.5 |
| 澳大利亚 | Burdekin Estuary | 617~13 031 | 221.0 |
| 澳大利亚 | Fitzroy Estuary | 699~7 947 | 139.2 |
| 澳大利亚 | Johnstone Estuary | 387~9 744 | 110.6 |
| 澳大利亚 | Korogoro Creek | 869.0 | |
| 澳大利亚 | Evans Head | 997±317 | 63.0±166.0 |
| 澳大利亚 | Darwin | 622~1 263 | 40.0 |
| 澳大利亚 | Hinchinbrook Island | 1 341~3 304 | 30.0 |
| 澳大利亚 | Jacobs Well | 531~5 036 | 19.0 |
| 澳大利亚 | Newcastle | 404~3 224 | 46.0 |
| 澳大利亚 | Seventeen Seventy | 314~1 399 | 10.0 |
| 澳大利亚 | Barwon Heads | 0.4 | |
| 巴布亚新 几内亚 | Nagada Creek | 540~1 680 | 43.6 |
| 巴哈马 | Norman’s Pond | 395~690 | 13.8 |
| 巴西 | San Francisco Estuary | 298~2 007 | 0.7 |
| 巴西 | Itacuraca Creek | 660~7 700 | 113.5 |
| 百慕大群岛 | Mangrove Bay | 268~4 823 | 65.0 |
| 肯尼亚 | Tana River Delta | 230~5 300 | 58.0 |
| 肯尼亚 | Gazi Bay | 575~6 435 | 54.3 |
| 马达加斯加 | Betsiboka | 270~1 530 | 9.1±14.2 |
| 美国 | Shark River | 975~6 016 | 102.0 |
| 坦桑尼亚 | Ras Dege | 400~5 050 | 33.7 |
| 坦桑尼亚 | Mtoni | 400~1 700 | 18.0 |
| 印度 | Malta Estuary | 358~3 033 | 2.3 |
| 印度 | Dhamra Estuary | 146~751 | 0.6~218.0 |
| 印度 | Thakuran Estuary | 160~737 | 0.3 |
| 印度 | Sundarbans | 376~561 | 4.7 |
| 印度 | Gaderu Creek | 2 215±864 | 56.0±100.9 |
| 印度 | Gautami Godavari Estuary | 430~4 770 | 43.4 |
| 印度 | Mooriganga Estuary | 152~1 530 | 7.7 |
| 印度 | Saptamukhi Estuary | 193~4 000 | 28.5 |
| 印度 | Cochin | 1 500~3 800 | 55.1 |
| 印度 | Hooghly Estuary | 70~1 530 | -2.8~84.4 |
| 越南 | Kien Vang | 713~8 136 | 93.5 |
| 越南 | Tam Giang | 528~11 481 | 135.0 |
| 中国 | 漳江口云霄溪 | 1 733±605 | 57.6~115.2 |
| 中国 | 东寨港 | 426~8 910 | 12.1(港内+河道) |
| 中国 | 东寨港 | 396~8 910 | 6.1(整个区域) |
3.4.3 东寨港CO2排放对红树林植物固碳量的抵消作用
红树林碳汇可以用植物的年固碳量进行表征[27],以东寨港红树林面积为1 771 hm2,单位面积植物固碳量为6.55~13.74 t·hm-2·a-1[28]进行计算,其红树林植物固碳量为(1.16~2.43)×104 t。仅以东寨港港内水域与河流作为计算区,水域面积为39.74 km2,单位面积CO2通量为12.1 mmol·m-2·d-1,每年可以CO2向大气释放碳2.11×103 t,即抵消约红树林8.6%~18.2%的植物固碳量;以东寨港及周边水域(整个研究区域)作为计算区,水域面积为94.95 km2,单位面积CO2通量为6.1 mmol·m-2·d-1,则每年以CO2向大气释放碳2.54×103 t,即抵消约红树林10.4%~21.9%的植物固碳量。
4 结论
1)东寨港水体理化参数时空分布特征:盐度与pH呈现出港外最高,港内次之,河流最低的特征;pCO2大体上表现出与盐度、pH相反的空间分布特征。温度与pCO2为干季低于湿季,DO则是干季高于湿季,盐度的干湿季差异不明显。
2)温度、物理混合与生物活动共同调控东寨港pCO2的时空分布。尽管5月份河口水体存在净光合作用,但是表层水中的pCO2仍然高于大气水平。
3)河流是CO2的强源,湿季CO2通量是干季的6.2倍;东寨港及周边水域CO2通量干季平均值为3.4±3.6 mmol·m-2·d-1,湿季为8.8±8.2 mmol·m-2·d-1,年平均为6.1±6.3 mmol·m-2·d-1,在全球红树林河口中处于较低水平,每年的CO2释放量将抵消红树林10.4%~21.9%的植物固碳量。